- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT Resting-state functional connectivity (RSFC), the temporal correlation of intrinsic activation between different brain regions, has become one of the most fascinating field in the
functional imaging studies. To better understand the association between RSFC and individual creativity, we used RSFC and the figure Torrance Tests of Creative Thinking (TTCT-F) to
investigate the relationship between creativity measured by TTCT and RSFC within two different brain networks, default mode network and the cognitive control network, in a large healthy
sample (304). We took the medial prefrontal cortex (MPFC) and the bilateral dorsolateral prefrontal cortices (DLPFC) to be the seed regions and investigated the association across subjects
between the score of TTCT-F and the strength of RSFC between these seed regions and other voxels in the whole brain. Results revealed that the strength of RSFC with the MPFC was
significantly and negatively correlated with the score of TTCT-F in the precuneus. Meanwhile, we also found that the strength of RSFC with the left DLPFC was significantly and positively
correlated with the score of TTCT-F in the right DLPFC. It suggests that the decreased RSFC within DMN and the increased RSFC within CCN presents a potential interaction mechanism between
different region for higher creativity. SIMILAR CONTENT BEING VIEWED BY OTHERS THE INDISPENSABLE ROLE OF THE CEREBELLUM IN VISUAL DIVERGENT THINKING Article Open access 06 October 2020
STATIC AND DYNAMIC FUNCTIONAL CONNECTIVITY SUPPORTS THE CONFIGURATION OF BRAIN NETWORKS ASSOCIATED WITH CREATIVE COGNITION Article Open access 08 January 2021 DYNAMIC SWITCHING BETWEEN BRAIN
NETWORKS PREDICTS CREATIVE ABILITY Article Open access 15 January 2025 INTRODUCTION Creativity is necessary to the growth of human society, economy and social culture and has generally been
viewed as “a kind of novelty that is useful, valuable and generative”1. Divergent thinking has been regarded as a critical aspect of creativity2. The research of divergent thinking has a
long history and is the biggest research field in creativity study3,4. A recent meta-analysis5 indicated that divergent thinking is an valid predictor of creative ability. Divergent thinking
has been defined as the capacity to generate many possible solutions to a given problem3. Torrance Test of Creative Thinking (TTCT)6, which is based on divergent thinking, has an extensive
application in the test of creativity7 and is widely quoted of many creativity tests8. Most of previous studies mainly focus on the neural basis of verbal creativity, the mount of the study
of the neural basis of visual creativity is little. In addition, the verbal test would be more influenced by educational background and confused by intellectual capacity than figure
test9,10,11. What’s more, visual creativity, the production of novel and useful visual forms, is a primary component of fields such as painting, photography, sculpture and architecture12. To
date, no functional connectivity studies have been published in this domain of creativity research. The primary aim of this study was to investigate how visual creativity, as measured by
TTCT-F, correlates to functional connectivity derived from resting-state brain imaging. Previous studies have suggested that creativity requires interactions of multiple regions and
left-right brain and should thus not be ascribed to one particular hemisphere or region13. Researchers have long studied the biological basis for creativity and have found increasing
evidence relating high performance on creative tests to the coordination of multiple brain regions, utilizing both structural and functional brain imaging
techniques14,15,16,17,18,19,20,21,22. Communication between brain regions may be crucial in complex cognitive processes (e.g., creativity)23. Examination of resting-state functional
connectivity (RSFC), which reflects temporal correlations between blood oxygen level-dependent signals in different brain regions during rest, can indicate direct or indirect functional
relations between brain regions24,25. The researches of RSFC are quickly growth and become one of the hottest theme of human imaging study26. Region-of-interest (ROI) analysis is one of the
most commonly used methods to examine functional connectivity during rest24,27. Previous studies had demonstrated that RSFC for both the region of interest (ROI)-based has a high degree of
test-retest reliability28,29. Previous studies have consistently demonstrated that there are similar patterns of RSFC network in most cortical systems in the human brain, e.g. visual
network, sensorimotor network, default mode network (DMN), cognitive control network (CCN)24,30,31,32 and salience network (SN)33. Recent findings suggested that specific networks and brain
regions are related to individuals creativity and any single brain region can not achieve creativity13,34. In other words, creativity may be associated with the strength of RSFC among
specific brain regions and the interactions between these brain regions might be crucial. The DMN network was consistently suggested correlated with creativity, which mainly included: medial
prefrontal cortex (MPFC), posterior cingulate cortex (PCC)/precuneus and lateral temporal/parietal lobe. According to Randy Buckner and colleagues, the DMN network is involved in
“constructing dynamic mental simulations based on personal past experiences such as used during remembering, thinking about the future and generally when imagining alternative perspectives
and scenarios to the present.” A recent RSFC study found that higher creativity as measured by divergent thinking (DT) test, a verbal creativity test, was positively associated with the
strength of RSFC between the medial prefrontal cortex (MPFC) and the posterior cingulate cortex (PCC)14. Another research investigated the association between creativity as measured by
verbal TTCT and the RSFC in DMN and revealed that creativity was positively correlated with the strength of RSFC between the MPFC and the middle temporal gyrus (MTG)35. These results seem to
show verbal creativity was related to the consistency of spontaneous fluctuations among DMN regions. Additionally, DMN brain regions are often involved in creativity experiments. Previous
research found that MPFC was more active in the creative task not only in verbal divergent thinking task (creative story generation)36, but also in visual creativity12. Both MPFC and
PCC/precuneus were also more active in insightful problems solving37,38,39,40. Thus, the strength of RSFC in DMN may be crucial to the visual creativity as measured by figure TTCT. Previous
functional imaging studies have focused on the contribution of CCN, which mainly included dorsolateral PFC (DLPFC) and dorsal anterior cingulate cortex (dACC), to creative task. The brain
regions of CCN (e.g. DLPFC and dACC) were frequently activated by many kinds of creative tasks. For example, greater activation were found in creative story generation36, divergent
thinking41, improvisational music playing42 and insightful problems solving39,43. More importantly12, investigated brain activation during participants solving the visuo-spatial creativity
problem and found visual creativity task more activated DLPFC and MPFC relative to the control task. It is thus reasonable to speculate whether the strength of RSFC in CCN may be related to
visual creativity as measured by figure TTCT, in spite of undetected association between verbal creativity and the strength of RSFC with DLPFC14,35. Depending on these tasks, different brain
networks recruited in the creative cognition, such as DMN and CCN networks. It meant that communication between brain regions were critical to understanding the neuroscience of creativity.
However, no direct evidence has been provided from the perspective of brain networks. Our hypothesis, inspired by these earlier findings, is that higher creativity test scores may correspond
to more efficient information transfer in the brain. In particular the relationship between individual creativity and functional connectivity properties of the brain network has rarely been
investigated, leaving the impact of large-scale brain networks on creativity largely unknown. In the present study, we aimed to further investigate the association between visual creativity
as measured by figure TTCT and the strength of RSFC within DMN and CCN. We chose TTCT-F rather than verbal TTCT because the verbal test would be more influenced by educational background
and confused by intellectual capacity than figure test9,10,11. If as mentioned above, DMN and CCN brain regions were attribute to individuals creativity, we should observe a relationship
between verbal creativity and RSFC. RESULTS BEHAVIORAL RESULTS The mean and the standard deviation of TTCT-F, CRT scores and age are shown in Table 1. There were no significant correlations
between the psychological and demographic measures (CRT score, sex and age) and the total scores of TTCT-F. There were also no significant difference between males and females in the total
score of TTCT-F, each dimension score of TTCT-F and the score of CRT (the significance level in these analysis are bigger than 0.1). CORRELATION OF CREATIVITY WITH THE STRENGTH OF RSFC WITH
THE MPFC We examined brain regions that showed significant correlation between the total scores of TTCT-F and the strength of RSFC with the MPFC. After controlling the effects of age, sex,
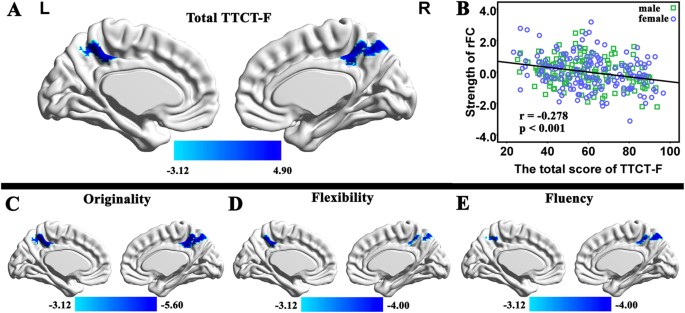
CRT scores and mean FD, the multiple regression analysis revealed that the TTCT-F total score was significantly negatively correlated with the strength of RSFC between the MPFC and the
precuneus [x, y, z = 0, −51, 48, _t_ = −4.78, 196 voxels, _p_ = 0.039 corrected using the whole-brain voxel-level FDR approach; see Fig. 1A]. No significant positive association was found
between the total TTCT-F score and the strength of RSFC with the MPFC. To verify the found of behavioral data, in the analysis of fMRI data, we also examined brain regions that showed
significant correlation between each dimension of creativity (originality, flexibility and fluency) and the strength of RSFC with the MPFC. Three whole-brain multiple regression analyses
were performed separately including the score of each dimension, age, sex, the CRT score and mean FD as covariates. Significant negative correlation were only observed in the analyses of
originality dimension (x, y, z = 0, −51, 51, _t_ = −5.58, _p_ = 0.001 corrected using the whole-brain voxel-level FDR approach). In the other analysis, we found similar tendencies for the
correlation between the dimensions of creativity (flexibility and fluency) and the strength of RSFC with the MPFC (statistical values and coordinates of the peak voxel in this region were as
follows: x, y, x = −3, –48, 48, t = −3.91 in the analysis of flexibility; x, y, x = 0, –48, 45, t = −3.99 in the analysis of fluency). The high consistency among the total TTCT-F score and
three dimensions of the TTCT indicated that each dimension could not provide meaningfully more information. CORRELATION OF CREATIVITY WITH THE STRENGTH OF RFC WITH THE BILATERAL DLPFCS We
examined brain regions that showed significant correlation between the total scores of TTCT-F and the strength of RSFC with the bilateral DLPFCs to determine whether creativity is also
associated with RSFC in networks other than DMN. After controlling the effects of age, sex, CRT scores and mean FD, the multiple regression analysis revealed that the TTCT-F total score was
significantly positively correlated with the strength of RSFC between the left DLPFC and the right DLPFC (x, y, z = 42, 39, 24, _t_ = 5.18, 69 voxels, _p_ = 0.006 corrected using the
whole-brain voxel-level FDR approach; see Fig. 2). Meanwhile significant positive correlation was found between the individual TTCT-F total score and the strength of RSFC between the right
DLPFC and the left DLPFC (x, y, z = −36, 45, 12, _t_ = 4.98, 4 voxels, _p_ = 0.031 corrected using the whole-brain voxel-level FDR approach; see Fig. 3). No significant negative association
was found between the total TTCT-F score and the strength of RSFC with the left DLPFC and no significant negative relationship between the total TTCT-F score and the strength of RSFC with
the right DLPFC. We also examined brain regions that showed significant correlation between each dimension of creativity (originality, flexibility and fluency) and the strength of RSFC with
the left DLPFC. Three whole-brain multiple regression analyses were performed separately including the score of each dimension, age, sex, the CRT score and mean FD as covariates. Significant
positive correlation were observed in the analyses of originality dimension (x, y, z = 42, 39, 24, _t_ = 5.16), fluency dimension (x, y, z = 42, 39, 24, _t_ = 4.59) and flexibility (x, y, x
= 42, 36, 27, t = 4.21). Then we examined brain regions that showed significant correlation between each dimension of creativity (originality, flexibility and fluency) and the strength of
RSFC with the right DLPFC. The results revealed a similar tendency for the score of each dimension of creativity as measured by TTCT-F and the total score of TTCT-F (see fig. 3). The high
consistency among the results of multiple regression analyses in total TTCT-F score and three dimensions of the TTCT were highly consistent. CORRELATION OF GENERAL INTELLIGENCE WITH THE
STRENGTH OF RSFC WITH THE MPFC We further investigated brain regions that showed significant correlation between the score of CRT and the strength of RSFC with the MPFC. No significant
correlations were found between the CRT scores and the strength of RSFC with the MPFC in any of the regions. CORRELATION OF GENERAL INTELLIGENCE WITH THE STRENGTH OF RSFC WITH THE BILATERAL
DLPFCS We further investigated brain regions that showed significant correlation between the score of CRT and the strength of RSFC with the DLPFC. No significant correlations were found
between the CRT scores and the strength of RSFC with the DLPFC in any of the regions. INTERACTION EFFECTS BETWEEN SEX AND CREATIVITY ON THE STRENGTH OF RFC WITH MPFC After controlling the
effects of age, sex, the CRT score and mean FD, the voxel-wise ANCOVA revealed no significant interaction effects between the total TTCT-F score and sex in terms of the strength of RSFC with
the MPFC. INTERACTION EFFECTS BETWEEN SEX AND CREATIVITY ON THE STRENGTH OF RFC WITH BILATERAL DLPFCS After controlling the effects of age, sex, the CRT score and mean FD, the ANCOVA
revealed no significant interaction effects between the total TTCT-F score and sex in terms of the strength of RSFC with the right DLPFC. Besides, no significant interaction effects were
observed with the left DLPFC. DISCUSSION In the current study, we used fMRI to explore the association between RSFC and visual creativity as measured by TTCT-F. To the best of our knowledge,
this is the first study to investigate the association between individual visual creativity measured TTCT-F and RSFC. Our results revealed that higher creativity is related to the decreased
RSFC between the MPFC and the precuneus and the increased RSFC between left DLPFC and right DLPFC. Taken together, the results suggest that the altered RSFC within DMN and CCN might be
critically involved in visual creativity. Additionally, these results are consistent with the recent functional imaging researches of creativity that the brain regions of the DMN and CCN
would be essential to visual creativity12,40,44. Previous studies had indicated that the MPFC is a common part of the creativity network, such as divergent thinking tasks commonly activated
the MPFC13. For instance, recent two studies suggest that the altered RSFC with default mode regions was related to individual differences in divergent thinking measured by S-A creativity
test14 and verbal TTCT35. Both studies employed functional connectivity analysis with the MPFC specified as a seed region of interest. In addition, previous researches indicate that DT tasks
frequently activated MPFC, such as creative story generation36, insight problems solving37,38,39,40 and visual creativity task12,44. To some extent, our results consistent with previous
study about the association between DMN and visual creativity12. In addition, the precuneus was suggested to be involved in a wide range of tasks, such as visuo-spatial imagery, episodic
memory retrieval and self-processing operations (for a review to see)45. These cognitive activity is considered to be associated with creativity. Further more, a recent study used cerebral
blood flow (CBF) during rest (rest-CBF) to examine the association between rest-CBF and individual creativity and revealed regional rest-CBF in the precuneus was negatively correlated with
individual creativity46. Another research found that the reduced task induced deactivation (TID) in the precuneus during 2-back task is related to the higher creativity measured by S-A
creativity test47. This reduced TID in the DMN was suggested to be related to the reduced RSFC within the DMN48. The association between visual creativity measured by TTCT-F and the reduced
RSFC between the MPFC and the precuneus, in the present study, may consistent with the suggestion that the higher creativity is expected to be associated with reduced RSFC with the MPFC14.
Regions within the DMN have been reported in several recent studies of creativity14,35,12,44,49. Previous researchers have speculated about the potential role of the DMN in creativity. For
example, DMN has been involved in the process of blind-variation and selective-retention16 and internally-directed attention during divergent thinking49. Other researchers employed
Resting-state functional connectivity (RSFC) to explore the relationship between divergent thinking and functional connectivity in default mode regions14,35,50. Taken together, they all used
verbal divergent thinking problem as the measure of creativity. Here, we extend this finding beyond the verbal creativity into visual creativity. The DLPFC, one key node of CCN, was
suggested to be recruited in top-down cognitive control, working memory, sustained attention, cognitive flexibility13,51 suggested that the DLPFC is critical for creativity thinking.
Furthermore, the DLPFC was more activated by a wide range of creative task, such as creative writing17, creative story generation36, divergent thinking41, improvisational music playing42,52
and insightful problems solving39,43. Additionally, more activation in DLPFC was found in visuo-spatial creativity problem rather than the control task12. Ellamil, _et al._44 found greater
recruitment of DLPFC during the evaluation of creative book cover design and Huang, _et al._53 investigated the different brain activity between creative and uncreative figural TTCT and
found increased activity in the DLPFC. In addition, higher intelligence was related to the increased RSFC between left DLPFC and right DLPFC. That is to say the higher correlation within the
bilateral DLPFC and the better psychometric intelligence. The associated between the higher creativity measured by TTCT-F and increased RSFC between the left DLPFC and the right DLPFC might
reflect the notion that the tighter RSFC the better performance. Of course there are some limitations in the current study. Firstly, just as previous researches14,46,54,55,56, we enrolled
young healthy college students which are better educated. Thus the sample was lack of other population samples with different level of education. Although the interpretations of the results
have more or less limitation as previous researches on creativity14,46,47, the enrolled high-level education sample also can ensure our research purpose. Secondly, there is also one
disadvantage in this study as was the case with previous studies14,22,35. We only reported findings related to some predefined seed regions and did not included other techniques of data
analyses such as regional homogeneity or ICA analyses. Finally, we did not combined verbal and visual creativity measures in one study. This caused a very limited and fragmentary picture of
potential neural mechanisms involved in creativity. In conclusion, the current study revealed that the higher creativity, as measured by TTCT-F test, is related to the decreased RSFC between
the MPFC and the precuneus and the increased RSFC between the left DLPFC and the right DLPFC, which are the nodes belong to the DMN and CCN. These results may indicate that the altered
functional connectivity in the brain is crucial to higher creativity. METHODS PARTICIPANTS Three hundred and fifteen university students (169 females; mean age = 19.95 years, standard
deviation (SD) = 1.17) from Southwest University, Chongqing, China, were enrolled in this study. These participants are the part of our ongoing project to examine the association between
brain imaging, creativity and mental health. All the subjects reported no history of neurological or psychiatric problems (e.g., epilepsy, traumatic brain injury, neurodegenerative disorders
and cerebro-vascular disease). Both behavioral and MRI protocols were approved by the local ethics committee of Southwest China University. Written informed consent was obtained from all
participants prior to the study, which was approved by the Institutional Human Participants Review Board of Southwest University Imaging Center for Brain Research. The methods were conducted
in accordance with approved guidelines. Three participants were excluded because their unfinished test of TTCT and three participants because their quit of assessment of general
intelligence. Another six participants were discarded from further analyses because of extraordinary scanner artifacts, abnormal brain structures (e.g., unusually large ventricles) or their
heads motion more than 3 mm of translation or 3 degrees of rotation in any direction. Thus, three hundred and four participants remained in the topological properties analysis. The last
remaining participants were 140 males (mean age = 20.20 years, SD = 1.21) and 164 females (mean age = 19.72 years, SD = 1.17). ASSESSMENT OF INDIVIDUAL TTCT The TTCT was developed by
Torrance in 199657 and is the most widely used method designed to measuring creativity58 using verbal test (TTCT-V), figural test (TTCT-F) and auditory tests57. Since the TTCT-V is
influenced by the effects of education9 and general intelligence10, TTCT-F was used in order to remove these influence9 in our study. The TTCT-F involves three types of activities. All the
participants answer the same questions with a ten-minute time limit for each activity. The first activity requires subjects to constructs a picture based on an ellipse or jellybean shape
provided on the page. The second activity requires subjects to use 10 incomplete figures to make an object or picture. The third activity requires subjects to draw as many as possible
pictures or objects on three pages of vertical lines. The TTCT-F provides a total score which comprised three components: fluency (the number of relevant responses, which is associated with
the ability to generate a number of pictures or objects), flexibility (the number of different categories of responses, which reflects the ability to shift between conceptual fields) and
originality (The number of infrequent ideas, which reflects the ability to produce uncommon or unique responses)57,59. Total TTCT-F score is the sum of the score of the above three
component. Scoring was performed by three separate raters who were all blind to the study. The inter-rater reliability for scoring of the TTCT-F was at an acceptable level (Cronbach’s Alpha
> 0.85). The current study limited the analysis to the total creativity scores and did not include each component score because of the high correlation between the total creativity score
and each component score (each correlation coefficient > 0.92) and between the each other component score (each correlation coefficient >0.78). Consistent with the previous study58
which explored the association between regional gray matter volume (rGMV) and the total score of the TTCT-F. Another study11 also investigated the association between regional white matter
volume (rWMV) of the corpus callosum (CC) and the total score of the TTCT-F. Thus, we only using total TTCT-F score in the graphic theory analysis. Additionally, We conducted another
multiple regression analyses for each component of TTCT-F to investigate whether difference with each component of TTCT-F and with total TTCT-F score. ASSESSMENT OF GENERAL INTELLIGENCE In
order to examine individual general intelligence, we used the Combined Raven’s Test (CRT) to measure participants’ general intelligence, which is a recognized intelligence test with a high
degree of reliability and validity60. The reliability coefficient was 0.9261. The CRT, which included the Raven’s standard progressive matrix (C, D, E sets) and Raven’s colored progressive
matrix (A, AB, B sets), consisted of 72 nonverbal items revised by the Psychology Department of East China Normal University in 1989. For each item, the participant is required to select the
missing piece of a 3 × 3 matrix from one of eight alternatives. The score of this psychometric test, which is used as an index of individual intelligence, is equal to the number of correct
answers given by participants within a 40-minute period. IMAGING DATA ACQUISITION All functional images were obtained from a 3-T Siemens Magnetom Trio scanner (Siemens Medical, Erlangen,
Germany) at the Brain Imaging Research Central in Southwest University, Chongqing, China. The whole-brain resting-state functional images were acquired using T2-weighted gradient echo planar
imaging (EPI) sequence: slices = 32, repetition time (TR)/echo time (TE) = 2000/30 ms, flip angle = 90 degrees, field of view (FOV) = 220 mm × 220 mm, thickness = 3 mm, slice gap = 1 mm,
matrix = 64 × 64, resulting in a voxel with 3.4 × 3.4 × 4 mm3. During the functional images acquisition, participants were asked to close eyes lightly and keep still as much as possible. The
scan lasted for 484 s and acquired 242 volumes in total for each subject. Additionally, high-resolution T1-weighted anatomical images were acquired for each participant (TR = 1900 ms; TE =
2.52 ms; inversion time = 900 ms; flip angle = 9 degrees; resolution matrix = 256 × 256; slices = 176; thickness = 1.0 mm; voxel size = 1 × 1 × 1 mm). BEHAVIORAL DATA ANALYSIS All the
behavioral data were analyzed using the statistical software SPSS 13.0 (SPSS Inc., Chicago, IL, USA). Pearson correlations were carried out to examine the relationships between: the total
score of TTCT-F and each dimension score; the total score of TTCT-F and age; and the total score of TTCT-F and the score of CRT. The differences in the total score of TTCT-F, each dimension
score of TTCT-F and the score of CRT between males and females were also computed. PREPROCESSING OF IMAGING DATA The resting-state functional MRI data were preprocessed using Data Processing
Assistant for Resting-State fMRI (DPARSF) software62,63 based on SPM8 (http://www.fil.ion.ucl.ac.uk/spm/). Image preprocessing included the following steps: transformation of the data from
DICOM to NIfTI, removal of the first 10 volumes from each subject’s functional data, slice timing to correct the differences in image acquisition time between slices, realignment to correct
the head movement artifacts, spatial normalization of the functional images (voxel size resampled to 3 × 3 × 3 mm) into the MNI space using an echo-planar imaging (EPI) template64. All
subjects’ head movements were restricted to less than 3 mm in translation and 3 degrees in rotation. The images were then spatially smoothed with an isotropic gaussian kernel (8 mm Full
Width Half Maximume, FWHM). The resulting images were linearly detrended and filtered with a band pass filter (0.01–0.08 Hz). The nuisance signals (head-motion profiles, white matter,
cerebrospinal fluid and global signal) were also extracted and regressed out to remove the potential impact of those physiological artifacts. According to recent research that higher-order
models demonstrate benefits in reducing movement artifacts65,66, the Friston 24-parameter model was used to regress out head movement artifacts from the realigned data. The residual effects
of motion was regressed out in group statistical analysis by including mean framewise displacement (FD) derived with Jenkinson’s relative root mean square (RMS) algorithm as a regressors of
no interest66,67. These preprocessing steps were followed by the standard protocol published by Yan and Zang63,66. FUNCTIONAL CONNECTIVITY ANALYSIS Voxel-wise functional connectivity
analysis was performed using REST toolkit68 which based on SPM8. Previous research14 have found the divergent thinking test (measured by S-A creativity test) was closely correlated with the
strength of RSFC between the medial PFC (MPFC) and PCC. Additionally, a recent study35 found creativity (measured by TTCT-V)was positively related with the strength of RSFC between the MPFC
and the left middle temporal gyrus (MTG). The key node of the default mode network (DMN) was used as seed region in the two researches. In current study, a ROI was defined as a sphere with a
6-mm radius centered at the MPFC (−1, 47,−4), as reported in previous studies14,25,35. Besides, the relationship between individual creativity (measured by TTCT-F) and RSFC with other ROI
were performed to determine whether creativity is also associated with RSFC in CCN. For this purpose, we investigated the networks seeded by the bilateral dorsolateral prefrontal cortices
(DLPFC, center at −32, 44, 16 and 44, 36, 20)33. We then investigated the association between individual creativity and the RSFC with ROI to ascertain whether creativity is also related to
the RSFC in network other than DMN and CCN. To do this we investigated the network involving the bilateral orbital fronto-insula (FI, center at 38, 26, −10 and −32, 24, −10)33. The
functional connectivity map were computed through correlating the averaged time series of seed region and the time series of other voxels in the whole brain. The resulting correlation
coefficient map was then converted into z-map by Fisher’s r-to-z transformation to improve the normality. STATISTICAL ANALYSIS In order to test the relationship between individual creativity
measured by TTCT-F and RSFC with the ROI. Individual z value maps were entered into the second-level whole-brain analysis. Multiple linear regression analysis was employed to identify brain
regions in which RSFC strength with the MPFC was significantly correlated with individual creativity measured by the TTCT-F (total creativity score). Previous studies have indicated that
some aspects of brain asymmetries interact with gender. For example, males have a greater structural asymmetry of the plenum temporal than females69 and male brain is more functionally
lateralized or asymmetric in visual and auditory areas than the female brain70,71. In addition, sex-related differences also exist at a microscopic level, involving differences in
connectivity, neuronal density or synaptic efficiency72. Thus, the effects of sex, age, the score of CRT and mean FD were included as regressors of no interest. In addition, another three
multiple regression analyses were performed separately to examine the correlation between each dimension of creativity (originality, flexibility, fluency) and the strength of RSFC between
the MPFC and other voxels in the whole brain. For each multiple regression analysis, three covariates (age, sex, the CRT score and mean FD) were included as regressors of no interest.
Subsequently, multiple regression analysis was performed to examine whether there was a correlation between individual general intelligence measured by the CRT and RSFC with the MPFC. This
enabled us to establish whether there was any overlap between the regions of interest (ROIs; i.e., between ROIs associated with creativity and RSFC and those associated with general
intelligence and RSFC). Sex, age, total score of TTCT-F and mean FDwere included as regressors of no interest. Gender differences in creativity have been widely studied in behavioral and
neuro-scientific investigations21,73. Behavioral studies on gender differences in creativity have been inconclusive thus far with half the investigations reporting no significant differences
while the other half are characterized by mixed findings that, on average, favor females74. Neuro-scientific investigations on gender differences in creative thinking are rare. As far as we
know, only two researchers have directly addressed this issue21,73. In order to extend recent observations of sex differences in creativity, we also examined whether the relationship
between RSFC with the MPFC and creativity measured by TTCT-F differed between gender. A voxel-wise analysis of covariance (ANCOVA) was performed using the full factorial option in SPM8, in
which sex was defined as a group factor. Four covariates (age, score of CRT, total score of TTCT-V and mean FD) were included in the model and were all interacted with sex using the
interactions option in SPM8. These interaction effects were assessed using t-contrasts. To perform the multiple comparisons correction, a voxel-level false discovery rates (FDR)75 _p_ <
0.05 was selected. Generally, FDR is designed to control the expected proportion of false discoveries among the discoveries. The threshold of 0.05 means that on the average there was no more
than 5% of the discoveries to be false discoveries76,77. ADDITIONAL INFORMATION HOW TO CITE THIS ARTICLE: Li, W. _et al._ The Association between Resting Functional Connectivity and Visual
Creativity. _Sci. Rep._ 6, 25395; doi: 10.1038/srep25395 (2016). REFERENCES * Stokes, P. D. [Novelty] _Encyclopedia of Creativity_ [ Runco, M. A. & Pritzker, S. R. (ed.)][297–304]
(Academic press, San Diego, 1999). * Guilford, J. P. The nature of human intelligence, (McGraw-Hill, New York, 1967). * Guilford, J. P. Creativity. Am. Psychol. 5, 444–454 (1950). Article
CAS PubMed Google Scholar * Weisberg, R. W. Creativity: Understanding innovation in problem solving, science, invention and the arts, (Wiley, Hoboken, NJ, 2006). * Kim, K. H.
Meta-Analyses of the Relationship of Creative Achievement to Both IQ and Divergent Thinking Test Scores. J. Creative Behav. 42, 106–130 (2008). Article Google Scholar * Torrance, E. P.
Torrance tests of creative thinking, (Personnel Press, Incorporated, 1968). * Davis, G. A. [Identifying creative students and measuring creativity] _Handbook of gifted education_ [(ed. N.
Colangelo & G. A. Davis )][269–281] (Viacom, Needham Heights, MA, 1997). * Lissitz, R. W. & Willhoft, J. L. A methodological study of the Torrance Tests of Creativity. J. Educ. Meas.
22, 1–11 (1985). Article Google Scholar * Bornstein, R. A., Suga, L. & Prifitera, A. Incidence of Verbal IQ-Performance IQ discrepancies at various levels of education. J. Clin.
Psychol. 43, 387–389 (1987). Article Google Scholar * Kershner, J. R. & Ledger, G. Effect of sex, intelligence and style of thinking on creativity: A comparison of gifted and average
IQ children. J. Pers. Soc. Psychol. 48, 1033–1040 (1985). Article Google Scholar * Moore, D. W. et al. Hemispheric connectivity and the visual–spatial divergent-thinking component of
creativity. Brain Cogn. 70, 267–272 (2009). Article PubMed Google Scholar * Aziz-Zadeh, L., Liew, S. L. & Dandekar, F. Exploring the neural correlates of visual creativity. Soc. Cogn.
Affect. Neur. 8, 475–480 (2012). Article Google Scholar * Dietrich, A. & Kanso, R. A review of EEG, ERP and neuroimaging studies of creativity and insight. Psychol. Bull. 136, 822–848
(2010). Article PubMed Google Scholar * Takeuchi, H. et al. The association between resting functional connectivity and creativity. Cereb. Cortex 22, 2921–2929 (2012). Article PubMed
Google Scholar * Fink, A. et al. Gray matter density in relation to different facets of verbal creativity. Brain. Sruct. Funct., 1–7 (2013). * Jung, R. E., Mead, B. S., Carrasco, J. &
Flores, R. A. The structure of creative cognition in the human brain. _Front. Hum. Neurosci._ 7, 330 (2013). PubMed Google Scholar * Shah, C. et al. Neural correlates of creative writing:
An fMRI Study. Hum. Brain Mapp. 34, 1088–1101 (2013). Article PubMed Google Scholar * Zhu, F., Zhang, Q. & Qiu, J. Relating Inter-Individual Differences in Verbal Creative Thinking to
Cerebral Structures: An Optimal Voxel-Based Morphometry Study. PLoS One 8, e79272 (2013). Article ADS CAS PubMed PubMed Central Google Scholar * Beaty, R. E. et al. Creativity and the
default network: A functional connectivity analysis of the creative brain at rest. Neuropsychologia 64, 92–98 (2014). Article PubMed PubMed Central Google Scholar * Benedek, M. et al.
To create or to recall? Neural mechanisms underlying the generation of creative new ideas. NeuroImage 88, 125–133 (2014). Article PubMed Google Scholar * Ryman, S. G. et al. Sex
differences in the relationship between white matter connectivity and creativity. NeuroImage 101, 380–389 (2014). Article PubMed Google Scholar * Li, W. et al. Brain Structure Links Trait
Creativity to Openness to Experience. Soc. Cogn. Affect. Neur. 10, 191–198 (2015). Article Google Scholar * van den Heuvel, O. A. et al. Frontal-striatal abnormalities underlying
behaviours in the compulsive-impulsive spectrum. J. Neurol. Sci. 289, 55–59 (2010). Article PubMed Google Scholar * Fox, M. D. & Raichle, M. E. Spontaneous fluctuations in brain
activity observed with functional magnetic resonance imaging. Nat. Rev. Neurosci. 8, 700–711 (2007). Article CAS PubMed Google Scholar * Fox, M. D. et al. The human brain is
intrinsically organized into dynamic, anticorrelated functional networks. Proc. Natl. Acad. Sci. USA 102, 9673–9678 (2005). Article ADS CAS PubMed PubMed Central Google Scholar *
Shannon, B. J. et al. Premotor functional connectivity predicts impulsivity in juvenile offenders. Proc. Natl. Acad. Sci. USA 27, 11241–11245 (2011). Article ADS Google Scholar *
Greicius, M. Resting-state functional connectivity in neuropsychiatric disorders. Curr. Opin. Neurol. 21, 424–430 (2008). Article PubMed Google Scholar * Zuo, X.-N. et al. Reliable
intrinsic connectivity networks: test–retest evaluation using ICA and dual regression approach. Neuroimage 49, 2163–2177 (2010). Article PubMed Google Scholar * Shehzad, Z. et al. The
Resting Brain: Unconstrained yet Reliable. Cereb. Cortex 19, 2209–2229 (2009). Article PubMed PubMed Central Google Scholar * Raichle, M. E. Two views of brain function. Trends Cogn.
Sci. 14, 180–190 (2010). Article PubMed Google Scholar * Smith, S. M. et al. Correspondence of the brain’s functional architecture during activation and rest. Proc. Natl. Acad. Sci. USA
31, 13040 –13045 (2009). Article ADS Google Scholar * Beckmann, C. F., DeLuca, M., Devlin, J. T. & Smith, S. M. Investigations into resting-state connectivity using independent
component analysis. Philos. T. Roy. Soc. B. 360, 1001–1013 (2005). Article Google Scholar * Seeley, W. W. et al. Dissociable Intrinsic Connectivity Networks for Salience Processing and
Executive Control. J. Neurosci. 27, 2349–2356 (2007). Article CAS PubMed PubMed Central Google Scholar * Sawyer, K. The cognitive neuroscience of creativity: A critical review.
Creativity Res. J. 23, 137–154 (2011). Article Google Scholar * Wei, D. et al. Increased resting functional connectivity of the medial prefrontal cortex in creativity by means of cognitive
stimulation. Cortex 51, 92–102 (2014). Article PubMed Google Scholar * Howard-Jones, P. A., Blakemore, S.-J., Samuel, E. A., Summers, I. R. & Claxton, G. Semantic divergence and
creative story generation: An fMRI investigation. Cognitive Brain Res. 25, 240–250 (2005). Article Google Scholar * Jung-Beeman, M. et al. Neural activity when people solve verbal problems
with insight. PLoS Biol. 2, e97 (2004). Article PubMed PubMed Central Google Scholar * Fink, A., Benedek, M., Grabner, R. H., Staudt, B. & Neubauer, A. C. Creativity meets
neuroscience: Experimental tasks for the neuroscientific study of creative thinking. Methods 42, 68–76 (2007). Article CAS PubMed Google Scholar * Subramaniam, K., Kounios, J., Parrish,
T. B. & Jung-Beeman, M. A Brain Mechanism for Facilitation of Insight by Positive Affect. J. Cogn. Neurosci. 21, 415–432 (2009). Article PubMed Google Scholar * Fink, A. et al.
Enhancing creativity by means of cognitive stimulation: Evidence from an fMRI study. NeuroImage 52, 1687–1695 (2010). Article PubMed Google Scholar * Carlsson, I., Wendt, P. E. &
Risberg, J. On the neurobiology of creativity. Differences in frontal activity between high and low creative subjects. Neuropsychologia 38, 873–885 (2000). Article CAS PubMed Google
Scholar * Bengtsson, S. L., Csíkszentmihályi, M. & Ullén, F. Cortical Regions Involved in the Generation of Musical Structures during Improvisation in Pianists. J. Cogn. Neurosci. 19,
830–842 (2007). Article PubMed Google Scholar * Kounios, J. et al. The origins of insight in resting-state brain activity. Neuropsychologia 46, 281–291 (2008). Article PubMed Google
Scholar * Ellamil, M., Dobson, C., Beeman, M. & Christoff, K. Evaluative and generative modes of thought during the creative process. NeuroImage 59, 1783–1794 (2012). Article PubMed
Google Scholar * Cavanna, A. E. & Trimble, M. R. The precuneus: a review of its functional anatomy and behavioural correlates. Brain 129, 564–583 (2006). Article PubMed Google Scholar
* Takeuchi, H. et al. Cerebral Blood Flow during Rest Associates with General Intelligence and Creativity. PLoS One 6, e25532 (2011). Article ADS CAS PubMed PubMed Central Google
Scholar * Takeuchi, H. et al. Failing to deactivate: The association between brain activity during a working memory task and creativity. NeuroImage 55, 681–687 (2011). Article PubMed
Google Scholar * Broyd, S. J. et al. Default-mode brain dysfunction in mental disorders: A systematic review. Neurosci. Biobehav. Rev. 33, 279–296 (2009). Article PubMed Google Scholar *
Benedek, M. et al. To create or to recall? Neural mechanisms underlying the generation of creative new ideas. Neuroimage 88, 125–133 (2014). Article PubMed Google Scholar * Beaty, R. E.
et al. Creativity and the default network: A functional connectivity analysis of the creative brain at rest. Neuropsychologia 64, 92–98 (2014). Article PubMed PubMed Central Google
Scholar * Dietrich, A. The cognitive neuroscience of creativity. Psychonomic Bulletin & Review 11, 1011–1026 (2004). Article Google Scholar * Friis-Olivarius, M., Wallentin, M.
& Vuust, P. [Improvisation: the neural foundation for creativity] _Proceedings of the seventh ACM conference on Creativity and cognition_ [411–412] (ACM, Berkeley, California, USA,
2009). * Huang, P. et al. Evidence for a left‐over‐right inhibitory mechanism during figural creative thinking in healthy nonartists. Hum. Brain Mapp. 10, 2724–2732 (2012). Google Scholar *
Jung, R. E. et al. Neuroanatomy of creativity. Hum. Brain Mapp. 31, 398–409 (2010). PubMed Google Scholar * Takeuchi, H. et al. Regional gray matter volume of dopaminergic system
associate with creativity: Evidence from voxel-based morphometry. NeuroImage 51, 578–585 (2010). Article PubMed Google Scholar * Takeuchi, H. et al. Regional gray matter density is
associated with achievement motivation: evidence from voxel-based morphometry. Brain. Sruct. Funct., 1–13 (2012). * Torrance, E. P. The Torrance Tests of Creative Thinking-Norms-Technical
Manual Research Edition-Verbal Tests, Forms A and B-Figural Tests, Forms A and B, (Personnel Press, Princeton, NJ, 1966). * Gansler, D. A. et al. Cortical morphology of visual creativity.
Neuropsychologia 49, 2527–2532 (2011). Article PubMed Google Scholar * Kim, K. H. Can We Trust Creativity Tests? A Review of the Torrance Tests of Creative Thinking (TTCT). Creativity
Res. J. 18, 3–14 (2006). Article Google Scholar * Tang, Y. Y., Rothbart, M. K. & Posner, M. I. Neural correlates of establishing, maintaining and switching brain states. Trends Cogn.
Sci. 6, 330–337 (2012). Article Google Scholar * Li, D. & Chen, G. Combined Reven’s teat (CRT) - Chinese revised version (in Chinese), (East China Normal University, Shanghai, 1989). *
Wang, D. et al. Revision on the Combined Raven’s Test for the Rural in China. Psychol. Sci. 5, 23–27 (1989). MathSciNet Google Scholar * Yan, C. G. & Zang, Y. F. DPARSF: a MATLAB
toolbox for “pipeline” data analysis of resting-state fMRI. Front. Syst. Neurosci. 4, 13 (2010). Google Scholar * Ashburner, J. & Friston, K. J. Nonlinear spatial normalization using
basis functions. Hum. Brain Mapp. 7, 254–266 (1999). Article CAS PubMed PubMed Central Google Scholar * Satterthwaite, T. D. et al. An improved framework for confound regression and
filtering for control of motion artifact in the preprocessing of resting-state functional connectivity data. NeuroImage 64, 240–256 (2013). Article PubMed Google Scholar * Yan, C.-G. et
al. A comprehensive assessment of regional variation in the impact of head micromovements on functional connectomics. NeuroImage 76, 183–201 (2013). Article PubMed Google Scholar * Power,
J. D., Barnes, K. A., Snyder, A. Z., Schlaggar, B. L. & Petersen, S. E. Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion.
NeuroImage 59, 2142–2154 (2012). Article PubMed Google Scholar * Song, X. W. et al. REST: a toolkit for resting-state functional magnetic resonance imaging data processing. PLoS One 6,
e25031 (2011). Article ADS CAS PubMed PubMed Central Google Scholar * Kulynych, J. J., Vladar, K., Jones, D. W. & Weinberger, D. R. Gender Differences in the Normal Lateralization
of the Supratemporal Cortex: MRI Surface-rendering Morphometry of Heschl’s Gyrus and the Planum Temporale. Cereb. Cortex 4, 107–118 (1994). Article CAS PubMed Google Scholar * Hiscock,
M., Inch, R., Jacek, C., Hiscock-Kalil, C. & Kalil, K. M. Is there a sex difference in human laterality? I. An exhaustive survey of auditory laterality studies from six neuropsychology
journals. J. Clin. Exp. Neuropsychol. 16, 423–435 (1994). Article CAS PubMed Google Scholar * Hiscock, M., Israelian, M., Inch, R., Jacek, C. & Hiscock-Kalil, C. Is there a sex
difference in human laterality? II. An exhaustive survey of visual laterality studies from six neuropsychology journals. J. Clin. Exp. Neuropsychol. 17, 590–610 (1995). Article CAS PubMed
Google Scholar * Frost, J. A. et al. Language processing is strongly left lateralized in both sexes: Evidence from functional MRI. Brain 122 199–208 (1999). Article PubMed Google
Scholar * Abraham, A., Thybusch, K., Pieritz, K. & Hermann, C. Gender differences in creative thinking: behavioral and fMRI findings. Brain Imaging Behav. 1–13 (2013). * Pagnani, A. In
Encyclopedia of Creativity 2nd edn, (eds Pritzker S. R. & Runco M. A. ) 551–557 (Academic Press, 2011). * Benjamini, Y. & Yekutieli, D. The control of the false discovery rate in
multiple testing under dependency. Annals of statistics 29, 1165–1188 (2001). Article MathSciNet MATH Google Scholar * Assaf, M. et al. Abnormal functional connectivity of default mode
sub-networks in autism spectrum disorder patients. NeuroImage 53, 247–256 (2010). Article PubMed Google Scholar * Laird, A. R. et al. Comparison of the disparity between Talairach and MNI
coordinates in functional neuroimaging data: Validation of the Lancaster transform. NeuroImage 51, 677–683 (2010). Article PubMed Google Scholar Download references ACKNOWLEDGEMENTS This
research was supported by grants from the National Natural Science Foundation of China (31070900; 31271087; 31470981), the Program for New Century Excellent Talents in University (2011) by
the Ministry of Education, the Fundamental Research Funds for the Central Universities (SWU1209101) and the Key Discipline Fund of National 211 Project (TR201208-1) and the Research Fund for
the Doctoral Program of Jining Medical University (JY2015BS03). AUTHOR INFORMATION Author notes * Li Wenfu and Yang Junyi contributed equally to this work. AUTHORS AND AFFILIATIONS * School
of Mental Health, Jining Medical University, Jining, 272067, China Wenfu Li & Gongying Li * School of psychology, Southwest University, Chongqing, 400715, China Junyi Yang, Qinglin
Zhang & Jiang Qiu Authors * Wenfu Li View author publications You can also search for this author inPubMed Google Scholar * Junyi Yang View author publications You can also search for
this author inPubMed Google Scholar * Qinglin Zhang View author publications You can also search for this author inPubMed Google Scholar * Gongying Li View author publications You can also
search for this author inPubMed Google Scholar * Jiang Qiu View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS W.L. conducted the experiment,
analyzed the results and wrote the article. J.Y. and G.L. analyzed the results. J.Q. and Q.Z. conceived the experiment. All authors reviewed the manuscript. ETHICS DECLARATIONS COMPETING
INTERESTS The authors declare no competing financial interests. RIGHTS AND PERMISSIONS This work is licensed under a Creative Commons Attribution 4.0 International License. The images or
other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the
Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit
http://creativecommons.org/licenses/by/4.0/ Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Li, W., Yang, J., Zhang, Q. _et al._ The Association between Resting Functional
Connectivity and Visual Creativity. _Sci Rep_ 6, 25395 (2016). https://doi.org/10.1038/srep25395 Download citation * Received: 08 October 2015 * Accepted: 18 April 2016 * Published: 03 May
2016 * DOI: https://doi.org/10.1038/srep25395 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not
currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative









