- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT Microbial bioactive natural products mediate ecologically beneficial functions to the producing strains, and have been widely used in clinic and agriculture with clearly defined
targets and underlying mechanisms. However, the physiological effects of their biosynthesis on the producing strains remain largely unknown. The antitumor ansamitocin P-3 (AP-3), produced by
_Actinosynnema pretiosum_ ATCC 31280, was found to repress the growth of the producing strain at high concentration and target the FtsZ protein involved in cell division. Previous work
suggested the presence of additional cryptic targets of AP-3 in ATCC 31280. Herein we use chemoproteomic approach with an AP-3-derived photoaffinity probe to profile the proteome-wide
interactions of AP-3. AP-3 exhibits specific bindings to the seemingly unrelated deoxythymidine diphosphate glucose-4,6-dehydratase, aldehyde dehydrogenase, and flavin-dependent thymidylate
synthase, which are involved in cell wall assembly, central carbon metabolism and nucleotide biosynthesis, respectively. AP-3 functions as a non-competitive inhibitor of all three above
target proteins, generating physiological stress on the producing strain through interfering diverse metabolic pathways. Overexpression of these target proteins increases strain biomass and
markedly boosts AP-3 titers. This finding demonstrates that identification and engineering of cryptic targets of bioactive natural products can lead to in-depth understanding of microbial
physiology and improved product titers. SIMILAR CONTENT BEING VIEWED BY OTHERS ELICITING THE SILENT LUCENSOMYCIN BIOSYNTHETIC PATHWAY IN _STREPTOMYCES CYANOGENUS_ S136 VIA MANIPULATION OF
THE GLOBAL REGULATORY GENE _ADPA_ Article Open access 10 February 2021 UNLOCKING HIDDEN BIOACTIVE COMPOUNDS: FROM INDOLOCARBAZOLE AND RIPP BIOSYNTHESIS TO THE ACTIVATION OF CRYPTIC SECONDARY
METABOLISM VIA MICROBIAL INTERACTIONS Article 16 May 2025 BLOCKS IN THE PSEUDOURIDIMYCIN PATHWAY UNLOCK HIDDEN METABOLITES IN THE _STREPTOMYCES_ PRODUCER STRAIN Article Open access 12 March
2021 INTRODUCTION Microbes, exemplified by actinobacteria and fungi, are rich repertoire of natural products with diverse structures and bioactivities, which are used as antibacterial,
antifungal, antitumor, immunosuppressive, cholesterol-lowering agents, and so on1. Due to the wide applications of microbial natural products in clinic and agriculture, their modes of action
and targeted cellular components have been well characterized in diverse pathogens and infected tissues. For example, the antibacterial penicillin targets transpeptidases, belonging to the
penicillin-binding proteins (PBPs), and inhibits the peptidoglycan biosynthesis in bacteria2; The immunosuppressant rapamycin binds to the human FK506 binding protein FKBP12, and
subsequently the formed binary complex inhibits the target of rapamycin (TOR) protein to exert the immunosuppressive effect3. Interestingly, owing to the development of chemoproteomic
technology using drug-derived chemical probes, more drug binding proteins involved in multiple physiological pathways have been identified, implying additional biological functions of a
designated drug4. Using synthetic β-lactam derivatives with different side chain substitutions as probes for in vivo labeling, Staub and Sieber identified not only the known PBPs, but also
non-PBP bacterial targets, including the virulence factor ClpP and a resistance-related β-lactamase5. Recently, Sun et al. identified STAT3 as a new target of rapamycin and its suppression
of tumor growth, using a photoactive rapamycin analog6. However, the effects of natural product production upon their producing hosts have been underexplored. The cellular targets of a few
anti-infectives were largely deduced from the known target proteins in pathogens, and corresponding resistant mechanisms were characterized in the producing strains7. As the target of the
anti-tuberculosis rifamycin is the β-subunit of bacterial RNA polymerase8, the counterpart of which with mutations of N447, D438, and Q432 was identified in the rifamycin-producing strain
_Amycolatopsis mediterranei_ S699; and these mutants confer self-protection to the host9. Similarly, a designated 23S rRNA _N_-methyltransferase gene _lmrB_ was found to be present in the
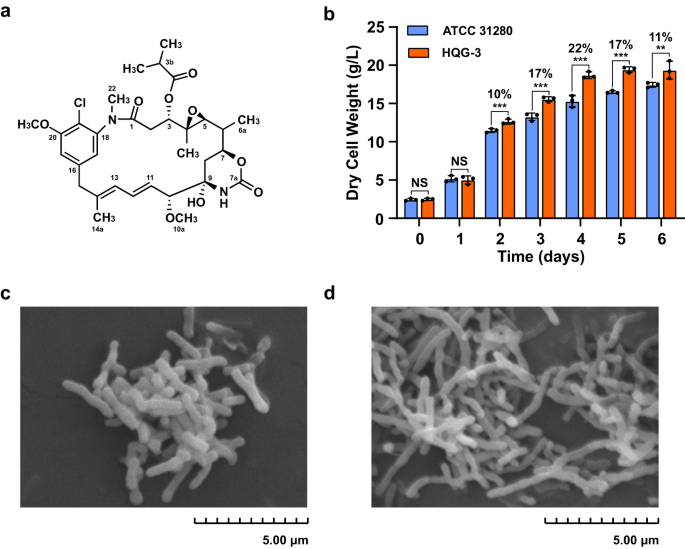
lincomycin biosynthetic gene cluster of _Streptomyces lincolnensis_ to provide resistance to supplemented lincomycin10. Ansamitocins (Fig. 1a) are actinobacteria-produced maytansinoids with
extraordinary antitumor potency; they specifically target microtubules and have served as the “warheads” in immunoconjugates for the treatment of various types of metastatic breast cancers.
Ansamitocins are produced by _Actinosynnema_, _Amycolatopsis_ and _Nocardiopsis_, among which the members of the genus _Actinosynnema_ are mainly used for industrial fermentation of
ansamitocins11,12,13. Recently, we found that ansamitocin P-3 (AP-3) at a high concentration suppressed the growth of the producing strain, _Actinosynnema pretiosum_ ATCC 31280 (hereafter as
ATCC 31280) (Supplementary Table 1), by targeting the cell division protein FtsZ, a β-tubulin-like protein in bacteria. Overexpression of the FtsZ-encoding gene obviously improved the
resistance of the recombinant strain to AP-3, as evidenced by a better growth than the starting strain in the presence of 300 mg/L AP-3. However, growth of the recombinant strain still
suffered inferior biomass in the presence of exogenous AP-3, suggesting that FtsZ might not be the only target of AP-314. Therefore, we proposed that other cryptic target(s) of AP-3 might be
present in the producing strain, and identification of these targets would be crucial to understand the stress caused by AP-3 biosynthesis and then further improve the AP-3 titer. Thus, we
used a specific photoaffinity AP-3 derived probe, combined with mass spectrometry (MS)-based quantitative analysis, to screen the cryptic targets of AP-3 in the producing strain ATCC 31280.
Our analyses revealed that the biosynthesis of AP-3 interfered the physiology of ATCC 31280 in a multi-target manner, with deoxythymidine diphosphate glucose-4,6 dehydratase (dTGD), aldehyde
dehydrogenase (ALDH), and flavin-dependent thymidylate synthase (FDTS) as AP-3 targets. The recombinant strains that overexpressed these proteins exhibited increased biomass under AP-3
exposure and also remarkably higher titers of AP-3. RESULTS CHEMOPROTEOMICS AND MS-BASED IDENTIFICATION OF CRYPTIC TARGETS OF AP-3 IN PRODUCING STRAIN ATCC 31280 To explore the potential
ability of AP-3 (Fig. 1a) to cause physiological perturbations of the producing strain, we disrupted the whole ansamitocin biosynthetic gene cluster (_ansa_-cluster) through double crossover
recombination in strain ATCC 31280 using pJTU1278-derived plasmid (Supplementary Table 2). This was achieved by cloning PCR-amplified upstream and downstream flanking sequences with two
pairs of designated primers (Supplementary Table 3), generating the recombinant strain HQG-3 as a reference strain (Supplementary Fig. 1). The growth of the two strains was compared in YMS
medium (Supplementary Table 4), and the results demonstrated that the biomass of HQG-3 was superior to that of strain ATCC 31280. The differences in biomass gradually increased during
exponential growth, so that the biomass of HQG-3 was 22% greater than that of the wild-type strain by the fourth day of fermentation (Fig. 1b), likely due to the absence of AP-3 production
in HQG-3. Observations on mycelial morphology in stationary phase by scanning electron microscopy revealed that strain ATCC 31280 had a morphology of short rods following fragmentation15,
while the mycelia of HQG-3 were branched and partially fractured (Fig. 1c, d). These results suggested that the biosynthesis of AP-3 at its low concentration also incurs physiological
stress, causing the inferior growth of the producing strain. To identify potential binding targets of AP-3 within strain ATCC 31280, we designed a specific chemoproteomic probe and performed
MS-based quantitative analysis. According to the structure-activity relationship of AP-3, the methylation at C20 has a negligible contribution to its biological activity13. Consequently, we
reasoned that this methyl group could be replaced by a diazirine-alkyne moiety (YNE) to generate a probe. Therefore, we disrupted the methyltransferase-encoding gene _asm7_16 in strain ATCC
31280 to generate mutant HQG-1 (Supplementary Fig. 2a, b), which accumulated 20-demethyl-ansamitocin P-3 (QG-1), as determined by liquid chromatography (LC)-MS analysis (Supplementary Fig.
2c–f). The YNE moiety was installed onto the phenolic hydroxyl group at C20 of QG-1 via the application of K2CO3 to generate the photoaffinity probe QG-YNE (Fig. 2a). The chemical structure
of compound QG-YNE was confirmed by MS and nuclear magnetic resonance analysis (Supplementary Figs. 3–5). Bioactivity analysis using the AP-3 sensitive yeast _Filobasidium uniguttulatum_ as
the indicator strain17 demonstrated that probe QG-YNE inhibited the yeast growth at a comparable level to AP-3 (Fig. 2b), indicating that the YNE moiety attachment did not interfere with the
biological potency of QG-YNE. To identify the AP-3 binding proteins, cell lysates of strain ATCC 31280 were pre-incubated with QG-YNE for 2 h and then irradiated with UV light. Following a
click reaction with streptavidin magnetic beads, we carried out pull-down manipulations to capture and enrich proteins bound to QG-YNE. Beads were separated from the proteins by in situ
tryptic digestion, and the reactions were then labeled with isobaric mass-tagging reagents (TMT-126/127) containing unique reporters (Supplementary Fig. 6). Finally, peptide fragments were
analyzed by MS, and the obtained functional proteomic information was filtered by a factor >1 for the ratio of TMT-127/126. The recovered proteins (fragments) were compared with the
annotated proteins of strain ATCC 31280, and from three independent surveys of AP-3 binding proteins using gradually reduced UV exposure times, we obtained enrichment of 708, 490 and 83
candidates, respectively. The effects of varied UV irradiations on individual outcomes were reproducible, and Venn diagram analysis revealed 14 binding proteins that were common to the three
UV exposure times (Fig. 2c). In addition to methyltransferase Asm7, which methylates the ansamitocin biosynthetic intermediate16, the other 13 candidate proteins were associated with
diverse metabolic pathways, including aldehyde metabolism, cell wall synthesis, nucleic acid biosynthesis, and regulatory pathways (Supplementary Table 5). These results suggested that the
stress on growth and morphological changes brought by AP-3 biosynthesis could arise through different metabolic pathways. AP-3 TARGETED DIFFERENT PATHWAYS TO INTERFERE WITH THE GROWTH OF THE
PRODUCING STRAIN As most of the AP-3 binding proteins participate in essential metabolic processes, such as cell division, cell-wall assembly, nucleotide biosynthesis and central carbon
metabolism, it was not possible to use gene disruption to examine their possible impacts on the producing strain. Therefore, we designed a set of integrative overexpression plasmids
containing each of the binding protein genes cloned under the control of a strong constitutive promoter _kasO_p_*_18 (Supplementary Figs. 7, 8). Asm7 in this case was excluded as it is
responsible for the methylation at C20 position of AP-3, and _APASM_6814_ and _APASM_6815_ form a gene cassette for their co-expression. Therefore, only 12 overexpression plasmids were
constructed and introduced separately into strain ATCC 31280 (Supplementary Table 2). The resulting recombinant strains were assessed for their tolerance to AP-3 by supplementing the
fermentation cultures with a final concentration of 200 mg/L AP-3. The biomass was then evaluated on the fifth day, when the cultures reached stationary phase. Strains HQG-7, HQG-13 and
HQG-17, which overexpressed aldehyde dehydrogenase (ALDH) (coded by gene _APASM_1052_), flavin-dependent thymidylate synthase (FDTS) (coded by gene _APASM_5765_) and deoxythymidine
diphosphate glucose-4,6 dehydratase (dTGD) (coded by gene _APASM_6307_), respectively, exhibited better AP-3 tolerance than the wild-type strain among all 12 mutants carrying overexpression
plasmids (Fig. 2d). Strains HQG-7, HQG-13 and HQG-17 were further challenged using a higher concentration of AP-3 (400 mg/L), and quantitative analysis of the biomass on the fifth day of
fermentation revealed better growth of these strains, with significantly enhanced tolerance of AP-3 as determined by boosting of the biomass by 45%, 42% and 27%, respectively, in comparison
to that of ATCC 31280 (Fig. 2e). As deduced from the annotation and functional studies in other bacteria on these three proteins, _APASM_1052_-coded ALDH catalyzes the oxidation of
aldehydes; _APASM_5765_-coded FDTS participates in the synthesis of dTMP from dUMP; and _APASM_6307_-coded dTGD mediates dTDP-L-rhamnose formation involved in the synthesis of bacterial cell
wall and capsule (Supplementary Fig. 9)19, indicating the diverse metabolic processes impacted by AP-3. AP-3 INHIBITED THE CATALYTIC ABILITY OF DTGD TO SUPPLY DTDP-L-RHAMNOSE FOR CELL WALL
ASSEMBLY dTGD oxidizes the C4 hydroxyl group of D-glucose with NAD(H) as a cofactor, followed by dehydration, which results in the generation of the dTDP-L-rhamnose precursor
dTDP-6-deoxy-D-_xylo_-4-hexulose (Supplementary Fig. 10). Various genetic studies have determined that abrogation of the dTDP-L-rhamnose biosynthesis greatly attenuates the virulence, and
can even influence the viability, of some pathogenic bacteria20. To examine the interaction between AP-3 and dTGD, we purified heterologously expressed dTGD from _E. coli_ (Supplementary
Fig. 11) and used surface plasmon resonance (SPR) to analyze the dTGD and AP-3 interaction at varied concentrations. AP-3 was determined to bind to dTGD with an affinity constant _K_D of
3.60 × 10–4 M, which was similar to the _K_D for the interaction between Asm7 from the AP-3 biosynthetic pathway16 and AP-3 (Fig. 3a, Supplementary Table 6). MS analysis of the dTGD peptide
fragments bound to the photoaffinity probe QG-YNE revealed that peptide 314-DNRDWWEPLKQR-325 was the specific incorporating peptide, which had the expected increase of 712.31 Da (equivalent
to QG-YNE with a loss of N2) in molecular weight, relative to the unmodified peptide fragment. Further analysis of the secondary MS (MS2) spectrum indicated that the labeling located to the
side chain carboxyl groups of Arg325 and Gln324 of dTGD (Fig. 3b, Supplementary Fig. 12). To gain a full understanding of the AP-3 interference with the catalytic ability of dTGD, the
molecular architecture of dTGD was established with the AlphaFold2 server. The flexible structure alignment of homologous proteins and DesIV from _Streptomyces venezuelae_ (PDB entry 1r66)
displayed that their architectures were highly similar with a RMSD of 0.41 and the conserved catalytic diad of Asp128 and Glu129 (Supplementary Fig. 13). Molecular docking analysis for AP-3
and dTGD suggested that AP-3 bound to the targeted protein with an affinity of −5.96 kcal/mol, through hydrogen-bond and hydrophobic interactions with adjacent amino acid residues in the
binding region. The binding area of AP-3 was away from the catalytic active site of dTGD, and the distances between the hydroxyl group at C20 of AP-3 and Asp128 and Glu129 of dTGD were 34.40
Å and 30.50 Å, respectively (Fig. 3c). The hydroxyl group at C20, which was modified to generate the photoaffinity probe QG-YNE, was near the Arg325 and Gln324 residues of dTGD, at a
distance appropriate for the photoaffinity group to crosslink with the residues, consistent with the MS2 analysis (Supplementary Fig. 14). Furthermore, we performed a quantitative analysis
of dTGD activity using different concentrations of dTDP-D-glucose as substrate and NAD+ as cofactor, as monitored by microplate reader at 320 nm (A320) (Supplementary Fig. 15). Under
optimized conditions, the _k_cat and _K_m of dTGD were measured as 43.29 min−1 and 0.15 mM, respectively. The _k_cat and _K_m values were also determined in the presence of 0.31 mM AP-3 as
34.46 min−1 and 0.17 mM, respectively, or in the presence of 0.63 mM AP-3 as 29.38 min−1 and 0.18 mM, respectively (Fig. 3d). The inhibitory constant (_K_I) of AP-3 was calculated as 1.10 mM
based on the associations between _K_m values and different concentrations of AP-3 (Supplementary Table 7). These results suggested that AP-3 serves as a non-competitive inhibitor of dTGD.
As disruption of the gene encoding dTDP-D-glucose 4,6-dehydratase resulted in the elongation of cells by up to 40-fold in _Proteus mirabilis_21, we compared the mycelial morphology of
strains ATCC 31280 and HQG-17. Interestingly, the average single-cell length of HQG-17 was about 1.65 times longer than that of ATCC 31280 statistically as observed by scanning electron
microscopy with stationary phase cultures (Fig. 3e), suggesting that AP-3 might inhibit dTDP-L-rhamnose supply in the producing strain by targeting dTGD. INTERFERENCE OF AP-3 WITH FDTS
ACTIVITY REDUCED THE BUILDING BLOCK SUPPLY FOR DNA SYNTHESIS The flavin-dependent thymidylate synthase (FDTS) executes a multi-step reaction pattern, which catalyzes the reductive
methylation of 2′-deoxyuridine-5′-monophosphate (dUMP) to 2′-deoxythymidine-5′-monophosphate (dTMP). During the methylation process, FDTS initially catalyzes the conversion of FAD to FADH2
with the consumption of two-equivalents of NADPH, and then transfers a methylene group from 5,10-methylenetetrahydrofolate (mTHF) to dUMP to generate dTMP and 7,8-dihydrofolate. Once the
mTHF is absent in the reactions under O2 existing conditions, FDTS acts as an NADPH oxidase and catalyzes FADH2 and two-equivalents of NADP to produce FAD and H2O2 (Supplementary Fig. 16)22.
To characterize the interaction between FDTS and AP-3, recombinant FDTS was obtained by heterologous expression in _E. coli_ (Supplementary Fig. 11). Based on SPR biosensor analysis, the
dissociation constant _K_D of AP-3 with FDTS was determined to be 6.65 × 10−5 M, indicating a high affinity of AP-3 for FDTS (Fig. 4a, Supplementary Table 6). To identify the residues
critical for AP-3 binding, QG-YNE-labeled FDTS was digested and analyzed by LC-MS/MS. Peptide 90-HFSYSQLSQR-99 was found to be covalently modified and exhibited a mass increase of 712.31
(equivalent to QG-YNE with loss of N2) compared to the unmodified peptide fragment. The amino acid sequence was confirmed unambiguously according to the numerous b-type ions formed after
high-energy collisional dissociation (HCD) fragmentation. Further analysis of the MS2 spectrum located the labeling to residue Leu96 (Fig. 4b, Supplementary Fig. 17). Subsequently, a
homo-tetramer structure of the FDTS was simulated with the AlphaFold2 server. The homologous proteins and ThyX from _Thermotoga maritima_ (PDB entry 7ndz) displayed high similarity with a
RMSD of 1.67 and the highly conserved Ser97 and Tyr99 in active sites (Supplementary Fig. 18). The molecular docking analysis showed that AP-3 bound to a region neighboring the catalytic
active pocket of FDTS with an affinity of −6.76 kcal/mol. AP-3 was anchored in the complex through hydrogen-bond and hydrophobic interactions with key amino residues. The distances between
the hydroxyl group at C20 of AP-3 and Ser97 and Tyr99 of FDTS were 11.70 Å and 12.40 Å, respectively (Fig. 4c). The hydroxyl group at C20 was closed to Leu96 with a 3.80 Å distance,
facilitating the photoaffinity group connected with the Leu96 during the UV irradiations (Supplementary Fig. 19). NADPH oxidase activity of FDTS was analyzed with FAD as the H+ acceptor and
in the presence of dUMP. Conveniently, NADPH was detectable through linear absorption at 340 nm in a range of 0.015 to 1 mM (Supplementary Fig. 20). The _k_cat and _K_m values of FDTS for
the reduction of NADPH were determined to be 57.63 min−1 and 0.66 mM, respectively, by monitoring the reactions with different concentrations of dUMP. As the catalytic ability of FDTS was
inhibited by AP-3 concentrations above 0.08 mM, reactions were supplemented with AP-3 at final concentrations of 0.02 or 0.04 mM. The _k_cat and _K_m values were further calculated as 42.88
min−1 and 0.65 mM in the presence of 0.02 mM AP-3, respectively, and 33.03 min−1 and 0.69 mM with 0.04 mM AP-3, respectively (Fig. 4d). These results showed that AP-3 was a non-competitive
repressor of FDTS, with a _K_I value of 0.05 mM (Supplementary Table 8). To further explore the physiological contribution of FDTS, we measured the intracellular dTMP concentrations from
strains ATCC 31280, HQG-3 and HQG-13. The concentration of dTMP in strain HQG-13 was increased by 14.3% and 8.3% over levels in strains ATCC 31280 and HQG-3, respectively (Fig. 4e,
Supplementary Fig. 21). Therefore, AP-3 production impedes the catalytic activity of FDTS and leads to a reduced dTMP supply for DNA synthesis in the producing strain. INTERACTION OF AP-3
WITH ALDH RESULTED IN REDUCED ACCUMULATION OF INTRACELLULAR ACETYL-COA ALDHs are commonly involved in the conversion of aldehydes, generated during cellular metabolic processes, into acids
with the assistance of cofactor NAD(P)+. Most aldehydes are chemically active and physiologically toxic for the generating organisms, and ALDHs participate in aldehyde detoxification and
assist in maintaining the reservoir of reducing equivalents23,24. To characterize the interactions between ALDH and AP-3, we purified soluble ALDH from the cell lysates of _Escherichia coli_
BL21(DE3) recombinants (Supplementary Fig. 11), and the affinity of AP-3 for ALDH was determined by SPR biosensor analysis. The _K_D value was determined to be 1.97 × 10−5 M for the
interaction of AP-3 with ALDH (Fig. 5a, Supplementary Table 6). To identify the ALDH residues bound by AP-3, recombinant ALDH was photo-labeled with QG-YNE, digested by trypsin and analyzed
via LC-MS/MS. The peptide 18-SRYDHFIGGEFTAPAK-33 exhibited a mass increase of 712.31 (equivalent to QG-YNE with loss of N2), and MS2 spectrum analysis of this peptide localized the labeling
to Arg19 (Fig. 5b, Supplementary Fig. 22). Then, a homo-tetramer architecture of the ALDH was constructed through the AlphaFold2 server. Comparison of homologous proteins and ALDH from
bovine mitochondria (PDB entry 1a4z) showed a high similarity with a RMSD of 0.82 and conserved Cys302 and Glu263 key residues in the catalytic active sites (Supplementary Fig. 23). Analysis
of the molecular docking complex revealed that AP-3 deviated from the catalytic active pocket of ALDH with an affinity of −7.40 kcal/mol. The distances between the hydroxyl group at C20 of
AP-3 and Cys302 and Glu263 of ALDH were 38.40 Å and 40.50 Å, respectively (Fig. 5c). Hydrogen-bond interactions were formed between the carbonyl group of the acetyl moiety at C3 and Arg17
and between the hydroxy group at C9 and Arg17, with the distances of 3.30 Å and 2.80 Å, respectively. The macrolactam ring of AP-3 is attached to the pocked of ALDH through hydrophobic
interactions. The hydroxyl group at C20 approached to Arg19 with a 4.20 Å distance, and such distance allowed a connection between the photoaffinity group of QG-YNE probe with Arg19 of ALDH
during the UV irradiations (Supplementary Fig. 24). The enzymatic reactions of ALDH were then established with aldehyde as the substrate and NAD+ as the cofactor. During the transformation
of aldehydes into acids by ALDH, the equivalent NADH was generated, which could be conveniently detected using a chromogenic assay (Supplementary Fig. 25). The _k_cat (8.40 min−1) and _K_m
(0.10 mM) values for the catalytic activity of ALDH on aldehydes were determined, and then AP-3 interference trials with ALDH were carried out. The _k_cat and _K_m values were calculated as
5.48 min−1 and 0.07 mM, respectively, with 0.31 mM of AP-3 in the reactions. _k_cat and _K_m values of 3.99 min−1 and 0.05 mM were further detected in the presence of 0.63 mM AP-3 (Fig. 5d).
These results suggested that AP-3 served as an inhibitor for ALDH, with an inhibition value of _K_I = 0.82 mM (Supplementary Table 9). Furthermore, overexpression of the ALDH-encoding gene
in strain HQG-7 enhanced the accumulation of intracellular acetyl-CoA by 90% over levels in strain ATCC 31280, and this increased acetyl-CoA consequently enhanced isocitric acid and citric
acid accumulation by 118% and 80%, respectively, in HQG-7 in comparison to that of strain ATCC 31280, boosting the growth of HQG-7 with ALDH overexpression. In addition, the intracellular
acetyl-CoA, citric acid and isocitric acid levels in the _ansa_-cluster-disrupted strain HQG-3 were also measured and found to be increased by 34%, 8% and 21%, respectively, in comparison to
that of strain ATCC 31280 (Fig. 5e, Supplementary Fig. 26, 27). AP-3 PRODUCTION BOOSTED BY TARGET ENGINEERING Since the strains that overexpressed the ALDH, FTDS or dTGD genes displayed
markedly increased biomass, when compared to the control strain, during the challenge with high concentrations of AP-3, we further examined the AP-3 production in these recombinants. Strain
ATCC 31280, HQG-7, HQG-13 and HQG-17 were cultured in YMS medium, and then AP-3 was extracted from the culture broths. AP-3 titers were determined to be 45.03 ± 0.95, 50.33 ± 5.46, 71.95 ±
6.95 mg/L and 83.47 ± 4.83 mg/L for strains ATCC 31280, HQG-7, HQG-13 and HQG-17, respectively, i.e., 11.77%, 59.78% and 85.37% AP-3 titer increases by the respective overexpression of ALDH,
FDTS and dTGD (Fig. 6a, Supplementary Fig. 28). Given the increased AP-3 titers by overexpression of targets, the overexpression constructs _APASM_5765_ (FDTS), _APASM_6307_ (dTGD) and
_APASM_1052_ (ALDH) were introduced into an AP-3 high-yielding strain WXR-24 and generated mutants HQG-23, HQG-24 and HQG-25, respectively. After fermentation in YMV medium, AP-3 titers were
measured and found to be 225 ± 33.25, 395.5 ± 31.02, 348.33 ± 22.49 and 254 ± 22.1 mg/L for WXR-24, HQG-23, HQG-24 and HQG-25, respectively (Fig. 6b). DISCUSSION Microbial bioactive natural
products mediate ecological beneficial functions and can confer evolutionary advantages over rival species25. For example, thiopeptides act as specific modulators of microbial phenotypes to
trigger biofilm formation26; at sub-inhibitory concentrations, tobramycin, tetracycline and norfloxacin function as signaling molecules to monitor the homeostasis of microbial communities
and can induce production of bioactive compounds to inhibit protozoan grazing27. However, the effects of natural product biosynthesis on their producing strains—be they beneficial or
deleterious—seem to be largely unknown. In our work, we confirmed the biosynthesis of ansamitocins incurred stress with reduced biomass to the producing strain, as shown either by
supplementation of exogenous AP-3 at high concentrations to the cultures14 or by deleting the ansamitocin gene cluster in the wild-type (Fig. 2b). Furthermore, we did systematic
investigation by structure simulation and binding protein capture with AP-3 derived photoaffinity probe, and identified FtsZ, the cytokinetic Z-ring protein essential for cell division14,
and deoxythymidine diphosphate glucose-4,6 dehydratase (dTGD), flavin-dependent thymidylate synthase (FTDS) and aldehyde dehydrogenase (ALDH) as the indigenous targets of AP-3, which are
largely responsible for the stress caused by AP-3 biosynthesis. Similar stress effects were observed in opportunistic pathogen _Pseudomonas aeruginosa_28 and marine bacterium _Phaeobacter
inhibens_29. The pyocyanin-producing wild-type _P. aeruginosa_ PA14 had a higher cell death ratio when compared to the Δ_phz_ mutant with pyocyanin biosynthetic gene cluster deleted, and
exogenous supplementation of pyocyanin increased the cell death ratio of the Δ_phz_ mutant28. In _Phaeobacter inhibens_, inactivation of the tropodithietic acid biosynthetic genes led to a
higher biomass and a higher carbon usage efficiency of the mutant than the wild-type29. Subsequent transcriptomic and proteomic comparisons revealed upregulated expressions of branched-chain
amino transferase IlvE, several uptake systems for amino acids and inorganic nutrients, and some components of the respiratory chain30. Chemoproteomic approaches have been successfully used
to identify the binding proteins of microbiota primary metabolites (fatty acids, aromatic amino acid-derived metabolites, vitamins, bile acids etc.)31, mycolic acids in mycobacteria32, bile
acid in _Clostridiodes difficile_33, and heme in Gram-positive and Gram-negative bacteria34. To our knowledge, however, chemoproteomics has not previously been deployed to explore the
binding spectrum of bioactive compounds in the producers. Whereas the activity-based protein profiling (ABPP) methodology uses chemical probes to specifically label catalytically active
proteins35, the photoaffinity approach explores probes with photoreactive moieties, such as the diazrine group, to interact and form crosslinks with any specific binding proteins36. As shown
in our experiments, three crosslinking reactions for the QG-YNE probe and cellular proteins resulted in efficient and specific labeling with 14 shared captured proteins (Fig. 2c), among
which Asm7 is with catalytic activity toward chloro-proansamitocin16, and AP-3 probably exerts allosteric effects on dTGD, FTDS, and ALDH (Figs. 3d, 4d, 5d). Therefore, chemoproteomics is
indeed a powerful strategy for characterizing the proteome-wide interactions of bioactive compounds in their producers. We have developed many genetic strategies for AP-3 titer increase,
including alleviation of rate-limiting post-PKS biosynthetic steps37, attenuation of the mycelial fragmentation15, improved AP-3 exportation through overexpression of export genes38, and
strengthened resistance by overexpression of the target protein FtsZ14 with titer increases between 24.94% and 93% and up to 330.6 mg/L. In strains that overexpressed the AP-3 target
proteins, we found that titers of AP-3 were boosted up to 85% and 76% over levels in wild-type strain ATCC 31280 and high-yielding strain WXR-24, respectively (Fig. 6). This result indicated
that the expression of target proteins in antibiotic producing strains is necessary for a designated antibiotic, and should be considered for titer improvement. Interestingly, while dTGD
and FTDS overexpression substantially enhanced the production of AP-3, the overexpression of ALDH only led to promoted growth of the producing strain and had no effect on AP-3 production.
ALDH oxidizes acetaldehyde to acetic acid, which can be further transformed into acetyl-CoA. Acetyl-CoA is not only a key molecule in the central carbon metabolism of microbes39, but also
enzymatically converted to the malonyl-CoA extender units for AP-3 biosynthesis40. Notably, the intracellular concentration of acetyl-CoA was 90% higher in HQG-7 overexpressing ALDH, than
that in strain ATCC 31280 (Fig. 5e). To explore the utilization of acetyl-CoA, we further examined key intermediates in the TCA cycle and found that intracellular concentrations of isocitric
acid and citric acid were dramatically enhanced by 118% and 80%, respectively, in HQG-7 when compared with strain ATCC 31280. Therefore, the increased acetyl-CoA flux in the ALDH
overexpression mutant is most likely diverted to the TCA cycle and biomass accumulation, rather than the biosynthesis of AP-3. In summary, our current research concentrated on the mining of
cryptic targets of the bioactive secondary metabolite AP-3 in its naturally producing strain through the power of chemoproteomics and an MS-based quantitative approach. AP-3 was found to
specifically bind with three protein targets and functioned as their non-competitive inhibitor. Our findings further suggested that AP-3 production causes a physiological stress on the
producer, which could be partly relieved by overexpression of the target proteins. Additionally, enhancing the expression of these targets also achieved titer improvement of AP-3. This
integration of target screening and engineering exhibits great potential for illuminating the biological effects of bioactive secondary metabolites and their cryptic targets, and for
improving strains for industrial application, including the production of antibiotics. METHODS STRAINS, PLASMIDS AND MEDIA Detailed information on culture media, strains, plasmids and
primers is provided in Supplementary Tables 1–4. _Escherichia coli_ strains DH10B, ET12567(pUZ8002) and BL21(DE3) were used as hosts for cloning, _E. coli-Actinosynnema_ bi-parental
conjugation and protein expression, respectively. Plasmid pJTU1278 was used to construct the gene disruption mutant. The integrative vector pLQ648, containing the constitutive strong
promoter _kasO_p*, was adapted to overexpress target protein-encoding genes, and pET30a was used for protein expression. During fermentation, strain ATCC 31280 and recombinants were firstly
cultured on YMG plates for 48 h at 30 °C, then the mycelia were transferred into S1 medium at 30 °C with shaking at 220 r.p.m. and cultured for 24 h. Subsequently, S1 culture was inoculated
into S2 medium at a final concentration of 3.3% (v/v) and cultivated at 30 °C and 220 r.p.m. for another 24 h. Finally, S2 seed broth was transferred into YMS medium at a final concentration
of 10% (v/v) and cultivated at 25 °C and 220 r.p.m. for 7 days to detect the biomass and AP-3 titer estimation in YMS/YMV media. QUANTITATIVE ANALYSIS OF AP-3 PRODUCTION BY LIQUID
CHROMATOGRAPHY-MASS SPECTROMETRY (LC-MS) For quantitative analysis of AP-3 (Supplementary Fig. 28) titers, an equal volume of methanol was supplemented into the final fermentation cultures
and stirred. The mixture was sonicated for 30 min. Then, 1 mL of the mixture was collected and centrifuged, and the supernatant was filtered through a 0.22 µm filter and subjected to LC-MS
analysis. For LC-MS determination, samples were analyzed on an Agilent series 1290-MS 6230 system (Agilent Technologies, Santa Clara, CA, USA) equipped with an Agilent Eclipse Plus C18
column (4.6 × 250 mm, 5 µm), with elution using 45% A (H2O with 0.5% formic acid): 55% B (acetonitrile) for 35 min and detection at 254 nm. The ion source was an AJS ESI model with positive
polarity. The gas temperature, flow and nebulizer pressure were 325 °C, 8 L/min and 35 psi, respectively. The fragmentor, skimmer and OCT 1 RF Vpp were set at 175, 65 and 750 V,
respectively. All derivatives of AP-3 were detected or isolated by this method. The _m/z_ of related compounds were calculated by high-resolution mass spectra and listed in corresponding
legends of supplementary figure. SYNTHESIS OF AN AP-3-DERIVED PHOTOAFFINITY PROBE 20-Demethyl-ansamitocin P-3 (QG-1) (46.5 mg, 0.075 mmol) was dissolved in anhydrous dimethyl formamide (DMF)
(1.0 mL), and then K2CO3 (20 mg, 0.15 mmol) and diazirine-alkyne moiety (YNE41) (synthesized by Bioduro-Sundia, Shanghai, China, 3.1 mg, 0.0125 mmol) were successively added into the
solution. After 1 h of stirring at 50 °C, the reaction was detected by thin-layer chromatography with mobile phase using 95% dichloromethane and 5% methanol, and additional YNE (4.4 mg,
0.0178 mmol) was added to the reaction mixture followed by incubation at 50 °C for another 3 h in the dark. The mixture was washed with saturated Na2CO3 and further dealt with 1 mol HCl and
brine. The organic layer was dried by anhydrous Na2SO4 and then filtered and concentrated. The crude extract was purified by high-performance liquid chromatography (HPLC) to obtain the
desired product, QG-YNE (18 mg, 80.6% yield). 1H and 13C nuclear magnetic resonance (NMR) spectra were recorded with a Bruker Advance 400 MHz in CDCl3 (1H NMR, 400 MHz; 13C NMR, 100 MHz)
spectrometer using tetramethylsilane as internal standard. MASS SPECTROMETRY-BASED AFFINITY-BASED PROTEIN PROFILING The procedure for target protein capture consisted of harvesting the
mycelia of strain ATCC 31280 on the third day by centrifugation at 3488 × _g_ (Thermo, TX-750), followed by washing with 7% NaCl, suspending in PBS buffer (150 mM NaCl, 3 mM KCl, 8 mM
Na2HPO4, 2 mM KH2PO4, pH 7.2–7.4) and crushing with an ultrasonic processor. The 10 mM chemical probe (QG-YNE) was pre-incubated with 50 µL of 30 mg/mL cell lysate for 2 h and then
irradiated with UV light at 365 nm for another 20/ 10/ 5 mins on ice for the generation of the covalent binding between the diazirine group on the probe and the amino acid residues of the
targeted proteins. The probe-binding proteins underwent another click reaction with 0.5 mM biotin-N3 assisted by 0.1 mM Tris-hydroxypropyltriazolylmethylamine (THTPA), 1 mM CuSO4 and 1 mM
vitamin C sodium (NaVc). Then, 50 µL streptavidin-sepharose (GE Healthcare) beads were poured into each reaction sample and incubated for 16 h with continuous rotation at room temperature.
The beads were washed with 600 µL 0.1 M PBS, 0.5 M NaCl, 4.0 M urea, and 100 mM triethylamonium bicarbonate (TEAB) consecutively. The enriched proteins were subjected to reductive alkylation
with 200 µL 10 mM Tris(2-carboxyethyl)phosphine (TCEP) and 200 µL 55 mM iodoacetamide. Enriched proteins were digested with 0.25 µg trypsin (Promega) overnight at 37 °C. The digests of both
dead-YNE (diazirine-alkyne moiety, Supplementary Fig. 6) and QG-YNE treated-samples were labeled with TMT2−126 and TMT2−127 Isobaric Label Reagent (Thermo Scientific), respectively. The
labeled sample was added to a Thermo Acclaim PepMap RSLC analytical column (75 µM × 15 cm) and washed by 0.1% formic acid in H2O and acetonitrile in 0.3 μL/min. Then, the peptide fragments
were analyzed on a Thermo Orbitrap Fusion Lumos proteomic mass spectrometer (Thermo Scientific)42. GENE KNOCKOUT AND OVEREXPRESSION The _asm7_ knockout mutant HQG-1 was obtained from strain
ATCC 31280 using a homologous recombination approach43. The left and right homologous arms were amplified by PCR using primer pairs deasm7-L-F/R and deasm7-R-F/R, respectively. After
sequencing validation, the DNA fragments were cloned into a pJTU1278-derived shuttle vector at the _Bam_HI/_Eco_RI and _Eco_RI/_Hin_dIII sites using a DNA Ligation Kit (TaKaRa). The
constructed plasmids were sequenced, introduced first into _E. coli_ ET12567(pUZ8002) by transformation and then into strain ATCC 31280 by conjugation. After culturing as a lawn of YMG with
0.2 mg/mL nalidixic acid and thiostrepton for 7 days at 30 °C, the exconjugants were further cultured without thiostrepton for two rounds on YMG solid medium. Then, the double crossover
mutants were screened by PCR to identify the target recombinants. The _asm7_ deletion mutant was named HQG-1, and the _asmA_ disruption recombinant HQG-3 was obtained in the same way as
HQG-1. Genes coding for the target proteins were overexpressed using a pSET152-derived integrative vector containing the strong constitutive promoter _kasO_p*. These genes were amplified by
PCR using corresponding primer pairs (Supplementary Table 3), and after sequencing validation, the target genes were individually inserted into the pSET152-derived shuttle vector at the
_Nde_I/_Eco_RI sites using the DNA Ligation Kit. The constructed plasmids were first introduced into _E. coli_ ET12567(pUZ8002) by transformation and then into the producing strains ATCC
31280 or WXR-24 by conjugation. After culturing on the lawn of YMG with 0.2 mg/mL nalidixic acid and apramycin for 4 days at 30 °C, the exconjugants were transferred onto YMG plates
containing apramycin and nalidixic acid, cultured for 2 days and then validated by PCR. EVALUATION OF THE AP-3 TOLERANCE OF ATCC 31280 AND RECOMBINANT STRAINS To test AP-3 tolerance, the
strains overexpressing the target genes (Supplementary Table 1) and the control strain ATCC 31280 were cultivated on YMG solid media for 2 days. Following the successive cultivation of the
seed broth in S1 and S2 media, 0.15 mL of S2 mycelium cultures (diluted to similar numbers of colony-forming units by OD600 detection) were transferred into 24-deep-well plates containing a
total volume of 1.5 mL YMS medium with a final concentration of 200 mg/L AP-3. Comparison of the biomass of the recombinants and strain ATCC 31280 was carried out after 5 days of
fermentation. Evaluation of the tolerance of the related strains against AP-3 was determined by dry cell weight on day 5. Additionally, HQG-7, HQG-13, HQG-17 and strain ATCC 31280 were
fermented with a final concentration of 400 mg/L AP-3 in the fermentation shake-flasks to investigate AP-3 tolerance. PROTEIN EXPRESSION, PURIFICATION AND ENZYME INHIBITION ASSAYS Gene
_APASM_1052_, _APASM_3207_, _APASM_5765_ and _APASM_6307_ coding for ALDH, Asm7, FDTS and dTGD, respectively, in strain ATCC 31280 were amplified by PCR using primer pairs ALDH-F/R,
Asm7-F/R, FDTS-F/R and dTGD-F/R (Supplementary Table 3), respectively. The obtained PCR products were inserted into the _Eco_RI/_Hin_dIII sites of the vector pET30a for sequencing
validation, and the verified plasmids were introduced into _E. coli_ BL21 for protein expression. The heterologous expression strains were incubated at 37 °C and 220 r.p.m. to an OD600 of
0.8 and induced with 0.1 mM isopropyl β–D-1-thiogalactopyranoside (IPTG), then successively cultured for another 10 h at 16 °C. The cells were harvested by centrifugation at 3488 × _g_
(Thermo, TX-750), 4 °C for 15 min, and then the pellet was resuspended in the corresponding buffers: Tris-HCl buffer (300 mM NaCl, 25 Mm Tris, pH 8.0) for dTGD; HEPES buffer (pH 7.5) for
FDTS; and Tris-HCl buffer (pH 8.5) for ALDH. After sonication on ice for 10 min with 2 s at 3 s intervals, the cell debris was removed by centrifugation at 13,800 × _g_ (Thermo Scientific
IEC MicroCL 21/21 R). and maintained at 4 °C for 30 min. Then the supernatant was applied to a nickel-NTA affinity chromatography column and eluted with resuspension buffer containing a
gradient of 50–250 mM imidazole. The 250 mM imidazole eluent was ultra-concentrated to 500 µL, and then resuspension buffer was added to wash away the imidazole, followed by
ultra-concentration again to 500 µL. The purified proteins were verified by SDS-PAGE (Supplementary Fig. 11) and used to analyze protein-AP-3 interactions and enzymic catalytic reactions.
Enzyme concentration was determined spectrophotometrically at 280 nm using NanoDrop One (Thermo Scientific). The biochemical reactions of dTGD were performed at 30 °C for 60 min in 100 μL of
50 mM Tris-HCl buffer (pH 8.0) containing 0.5 μg dTGD and 0.5 mM NAD+ with different concentrations of dTDP-D-glucose as the substrate (0.1, 0.16, 0.2, 0.32, 0.48, 0.64 or 0.8 mM). The
reaction was terminated by addition of 10 μL of 1 M NaOH. The final product, dTDP-6-deoxy-D-_xylo_−4-hexulose, was detected using a microplate reader at a wavelength of 320 nm to monitor the
reaction44. The corresponding _K_m and _V_max values were calculated by measuring the initial reaction rates of dTGD under different concentrations of dTDP-D-glucose. Based on this analysis
platform, 0.31 and 0.63 mM AP-3 were supplemented into the reactions to calculate the inhibition constants (_K_I) of AP-3 for dTGD. The catalysis reactions of ALDH were carried out at 30 °C
for 60 min in 100 μL of 50 mM Tris-HCl buffer (pH 8.5) containing 2.5 μg ALDH, 100 mM KCl, and 2 mM NAD+ with 0.01, 0.02, 0.04, 0.08, 0.12, 0.24, 0.48 or 0.96 mM acetaldehyde as the
substrate. Then, 0.1 mM water-soluble tetrazole salt (WTS-8) and 1 μM 1-methoxy-5-methylphenazinium methyl sulfate (1-mPMS) were added to the mixture to complete the reactions. With the
electron trapping agent of 1-mPMS, WTS-8 can react with NADH to generate formazan. The concentration of formazan was quantified by a microplate reader at a wavelength of 450 nm (A450)45
(Supplementary Fig. 25). The reaction constants of _K_m and _V_max were assessed by the absorption of A450 catalyzed by ALDH for different concentrations of acetaldehyde. Additionally, 0.31
and 0.63 mM of AP-3 were added to the reactions to calculate the inhibition constants (_K_I) of AP-3 for ALDH. Reactions with FDTS were conducted at 30 °C for 25 min in 100 μL of 50 mM HEPES
buffer (pH 7.5) containing 0.5 μg FDTS, 1 mM MgCl2, 62.5 μM FAD, and 0.6 mM NADPH with 0.1, 0.25, 0.5, 0.75, 1, 2, 4 or 8 μM dUMP as the substrate. The reactions were stopped by addition of
10 μL of 600 μM formic acid. The reaction constants of _K_m and _V_max for FDTS were determined by the consumption of NADPH to NADP using A340nm detection46. Additionally, 0.02 and 0.04 mM
of AP-3 were added into the reactions to calculate the inhibition constants (_K_I) of AP-3 for FDTS. SURFACE PLASMON RESONANCE BIOSENSOR ANALYSIS Surface plasmon resonance biosensor analysis
system was performed with a Biacore TM 8 K (GE Healthcare) for monitoring the biomolecular interactions at 25 °C. The buffer containing the purified proteins was exchanged with PBS for
preventing interference to the detection signal during the test. Then, the proteins were diluted to a concentration of 10 µg/mL with 10 mM sodium acetate buffer (pH 4.0) and then immobilized
on the surface of CM5 sensor chips. AP-3 was dissolved in PBS buffer with 0.5% DMSO and diluted to gradient concentrations of 100, 50, 25, 12.5 and 6.25 μM. During the binding process, AP-3
solution flowed across the CM5 surface in PBS buffer at a flow rate of 30 μL · min−1 for 90 s. Then in the dissociation step, PBS buffer was passed across the surface of CM5 at a flow rate
of 30 μL · min−1 for 120 s to elute AP-3. For every purified protein, responses signals from experiments with gradient concentrations of AP-3 were recorded, and the affinity constant (_K_D)
was calculated by Biacore Insight Evaluation Software. MICROSCOPY OBSERVATION Cell morphology was characterized by scanning electron microscopy (HITACHI S340II). The harvested mycelia were
washed three times with 0.1 M PBS buffer and fixed in 2.5% glutaraldehyde (dissolved in PBS buffer) for 12 h. Fixed cells were dehydrated with gradient concentrations of 5%, 15%, 30%, 60%,
80%, 90%, 95% and 100% ethanol solution (mixed with PBS buffer), and ethanol was removed using an automatic critical point dryer (LEICA). Dried mycelia were fixed on the copper platform with
conductive adhesive for surface gold-plating in a vacuum coater (LEICA). The images were captured with 10 k-fold magnification under scanning electron microscopy. The specimen size and
height were set as 51 mm and 1 mm. IDENTIFICATION OF QG-YNE-BINDING PEPTIDES Ten micrograms of recombinant His-tagged proteins ALDH, FDTS and dTGD in 100 μL PBS buffer were incubated with 10
μM of QG-YNE at room temperature for 2 h, followed by 365 nm UV irradiation for 20 min on ice. QG-YNE-labeled proteins were separated by SDS-PAGE, further extracted and dissolved in 50 mM
NH4HCO3 buffer. Chemical modification by reducing with 20 mM dithiothreitol at 60 °C for 60 min and alkylating with 50 mM iodoacetamide at 30 °C for 30 min were performed to reduce the
disulfide bonds and generate an open conformation of the proteins47. Then, the samples were digested with 2 μg trypsin (Thermo Scientific) at 37 °C for 12 h to generate peptide fragments.
The peptide fragment samples were desalted with Ziptip desalting columns (Pierce) and resuspended in 10 μL ddH2O containing 0.1% formic acid after drying by evaporation. The final samples
were analyzed using a Thermo Easy nLC1200/Q Exactive plus proteomic mass spectrometer to identify the amino acid residue(s) bound by the photoaffinity probe. EXTRACTION AND DETECTION OF
INTRACELLULAR METABOLITES After three days of fermentation, mycelia were harvested, and the intracellular metabolites were quantitatively analyzed. Specifically, 25 mL of fermentation broth
was collected and centrifuged at 6200 × _g_ (Thermo, F15-6×100y). for 12 min. The mycelia were washed with 25 mL of PBS buffer three times. Then, the washed mycelia were resuspended in 25 mL
of distilled water and divided equally into 5 tubes, followed by centrifugation at 13,800 × _g_ (Thermo Scientific IEC MicroCL 21/21 R). for 5 min and removal of the supernatant. Then, 2.5
mL of extracting solution containing 45% acetonitrile, 45% methanol and 10% glacial acetic acid was added, and the cell pellet was resuspended and sonicated for 15 min. After two
extractions, the combined mixture was centrifuged at 13,800 × _g_ (Thermo Scientific IEC MicroCL 21/21 R). for 5 min, and the supernatant was filtered through a 0.22 µm filter and collected
for analysis of the intracellular metabolites. All steps above were performed at 4 °C or on ice. The quantitative analysis of acetyl-CoA was performed using an LC-MS 1260-QQQ 6470 (Agilent)
with an Eclipse-C18 (4.6 × 250 mm, 5 μm, Agilent) column at a flow rate of 0.5 mL/min with a 2 μL injection volume. The mobile phase was composed of (A) 20 mM ammonium acetate in Milli-Q
water with a pH of 7.4 and (B) methanol. The metabolites were eluted by gradient mobile phase as follows: 0–5 min, 2% B; 5–15 min, 2% B to 50% B; 15–20 min, 90% B; 20–25 min, 90% B to 2% B;
and 25–30 min, 2% B. Quantitative analysis of citric acid, isocitric acid and dTMP was conducted using an ACQUITY UPLC H-Class system & Xevo TQ-XS triple quadrupole mass spectrometer
(Waters, Milford, MA, USA) with ACE Excel C18-PFP (100 × 2.1 mm, 1.7 μm, Phenomenex) at a flow rate of 0.3 mL/min with 1 μL injection volume. Water and acetonitrile, both containing 0.1%
formic acid, were used as mobile phases A and B, respectively, with a linear gradient of 1–100% B within 10 min. Metabolites were scanned by multiple reaction monitoring (MRM) mode with a
rate of 0.01 s/scan and a capillary voltage set at 1 kV. The source temperature was held at 150 °C while that of the desolvation gas was 450 °C. The flow rates of the desolvation gas
(nitrogen) and cone gas (argon) were 900 and 50 L/h, respectively. Acetyl coenzyme A was detected in positive ion and MRM modes. dTMP, citric acid and isocitric acid were detected in
negative ion and MRM modes. The calibration curves and MRM quantitative signal for quantification of dTMP are shown in Supplementary Fig. 21; for acetyl-CoA, in Supplementary Fig. 26; for
citric acid and isocitric acid, in Supplementary Fig. 27. The cone voltage (V) and collision energy (eV) of acetyl-CoA, citric acid, isocitric acid and dTMP were set as 20/15, 20/15, 20/20
and 15/20, respectively. EVALUATION OF BINDING MODELS FOR AP-3 AND TARGET PROTEINS Protein modeling was performed by AlphaFold v2.1.148 and ranked_0.pdb, which contained the prediction with
the highest confidence and selected for further analysis. All proteins were modeled in both the monomer format and homo-multimer format. To determine oligo states, amino sequences of ALDH,
FDTS and dTGD were used to search the PDB database (https://www.rcsb.org/), and homologous proteins were analyzed. ALDH was aligned to 5gtl with a sequence identity of 43%, which forms a
homo-tetramer. FDTS and dTGD were aligned to 3hzg and 1r66, which form a homo-tetramer and homo-dimer, respectively, with sequence identities of 72% and 70%, respectively. Protein structures
were analyzed by PyMol (Version 2.3.0)49. The molecular structure of AP-3 was obtained from the PDB database (PDB ID: 7e4p). AutoDock Tools50 was employed to convert protein and molecular
files into pdbqt with added polar hydrogen atoms. Molecular docking was performed by AutoDock Vina 1.2.351 with AP-3 and target proteins as the ligand and receptors, respectively. At the
same time, the argument exhaustiveness was set to 32 to give a more reliable docking result. The binding sites of AP-3 were visualized by PyMol (Version 2.3.0). The hydrogen bond
interactions and hydrophobic interactions between AP-3 and proteins were analyzed by LigPlot+ (Version v2.2.5)52 (Supplementary Figs. 14, 19 and 24). STATISTICS AND REPRODUCIBILITY One-way
ANOVA test was performed for the statistical analysis of biomass, mycelium length, AP-3 titer and concentrations of intracellular metabolites. _P_ ≥ 0.1, _P_ < 0.1, _P_ < 0.05, and
_P_ < 0.01 were defined as insignificant, significant, moderately significant, and highly significant, respectively. CONFIDENCE.T formula in Excel was used to estimate the 95%
confidence interval of mean. Cohen’s _d_ was employed to calculate the effect size of mean. Three independent biological replicates were used for the experiments of biomass measurement and
metabolites quantification, while five samples were chosen for the mycelium length measurement. The effects of AP-3 and probe QG-YNE on the growth rate of yeast indicator strain were
measured for one time. The interactions between AP-3 and deoxythymidine diphosphate glucose-4,6 dehydratase (dTGD), flavin-dependent thymidylate synthase (FDTS) and aldehyde dehydrogenase
(ALDH) were measured for one time. Three independent biological replicates were used in Michaelis-Menten constant measurement of three target proteins. REPORTING SUMMARY Further information
on research design is available in the Nature Portfolio Reporting Summary linked to this article. DATA AVAILABILITY The main data supporting the findings of this study are available within
the article and its Supplementary Information files. The datasets generated and analyzed during this paper are available from the corresponding authors upon request. The source data
underlying the graphs and proteomic charts are uploaded as a Supplementary Data 1 and 2. The sequences for all genes described in this manuscript are available in the GenBank databases under
the accession numbers CP073249.1. The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier
PXD043867. REFERENCES * Berdy, J. Bioactive microbial metabolites - a personal view. _J. Antibiot._ 58, 1–26 (2005). CAS Google Scholar * Izaki, K., Matsuhashi, M. & Strominger, J. L.
Glycopeptide transpeptidase and D-alanine carboxypeptidase: penicillin-sensitive enzymatic reactions. _Proc. Natl Acad. Sci. USA_ 55, 656–663 (1966). CAS PubMed PubMed Central Google
Scholar * Sabatini, D. M., Erdjument-Bromage, H., Lui, M., Tempst, P. & Snyder, S. H. RAFT1: a mammalian protein that binds to FKBP12 in a rapamycin-dependent fashion and is homologous
to yeast TORs. _Cell_ 78, 35–43 (1994). CAS PubMed Google Scholar * Drewes, G. & Knapp, S. Chemoproteomics and chemical probes for target discovery. _Trends Biotechnol._ 36, 1275–1286
(2018). CAS PubMed Google Scholar * Staub, I. & Sieber, S. A. Beta-lactams as selective chemical probes for the in vivo labeling of bacterial enzymes involved in cell wall
biosynthesis, antibiotic resistance, and virulence. _J. Am. Chem. Soc._ 130, 13400–13409 (2008). CAS PubMed Google Scholar * Sun, L. et al. Rapamycin targets STAT3 and impacts c-Myc to
suppress tumor growth. _Cell Chem. Biol._ 29, 373–385 (2022). CAS PubMed Google Scholar * Peterson, E. & Kaur, P. Antibiotic resistance mechanisms in bacteria: relationships between
resistance determinants of antibiotic producers, environmental bacteria, and clinical pathogens. _Front. Microbiol._ 9, 2928 (2018). PubMed PubMed Central Google Scholar * Ezekiel, D. H.
& Hutchins, J. E. Mutations affecting RNA polymerase associated with rifampicin resistance in _Escherichia coli_. _Nature_ 220, 276–277 (1968). CAS PubMed Google Scholar * Floss, H.
G. & Yu, T. W. Rifamycin-mode of action, resistance, and biosynthesis. _Chem. Rev._ 105, 621–632 (2005). CAS PubMed Google Scholar * Zhang, H. Z., Schmidt, H. & Piepersberg, W.
Molecular cloning and characterization of two lincomycin-resistance genes, _lmrA_ and _lmrB_, from _Streptomyces lincolnensis_ 78-11. _Mol. Microbiol._ 6, 2147–2157 (1992). CAS PubMed
Google Scholar * Prota, A. E. et al. A new tubulin-binding site and pharmacophore for microtubule-destabilizing anticancer drugs. _Proc. Natl Acad. Sci. USA_ 111, 13817–13821 (2014). CAS
PubMed PubMed Central Google Scholar * Venghateri, J. B., Gupta, T. K., Verma, P. J., Kunwar, A. & Panda, D. Ansamitocin P3 depolymerizes microtubules and induces apoptosis by binding
to tubulin at the vinblastine site. _Plos One_ 8, e75182 (2013). CAS PubMed PubMed Central Google Scholar * Cassady, J. M., Chan, K. K., Floss, H. G. & Leistner, E. Recent
developments in the maytansinoid antitumor agents. _Chem. Pharm. Bull._ 52, 1–26 (2004). CAS Google Scholar * Wang, X. R., Wang, R. F., Kang, Q. J. & Bai, L. Q. The antitumor agent
ansamitocin P-3 binds to cell division protein FtsZ in _Actinosynnema pretiosum_. _Biomolecules_ 10, 699 (2020). CAS PubMed PubMed Central Google Scholar * Wu, Y. T., Kang, Q. J., Zhang,
L. L. & Bai, L. Q. Subtilisin-involved morphology engineering for improved antibiotic production in actinomycetes. _Biomolecules_ 10, 851 (2020). CAS PubMed PubMed Central Google
Scholar * Spiteller, P. et al. The post-polyketide synthase modification steps in the biosynthesis of the antitumor agent ansamitocin by _Actinosynnema pretiosum_. _J. Am. Chem. Soc._ 125,
14236–14237 (2003). CAS PubMed Google Scholar * Tanida, S., Hasegawa, T., Hatano, K., Higashide, E. & Yoneda, M. Ansamitocins, maytansinoid antitumor antibiotics. producing organism,
fermentation, and antimicrobial activities. _J. Antibiot. (Tokyo)_ 33, 192–198 (1980). CAS PubMed Google Scholar * Wang, W. S. et al. An engineered strong promoter for Streptomycetes.
_Appl. Environ. Microbiol._ 79, 4484–4492 (2013). CAS PubMed PubMed Central Google Scholar * Kaminski, L. & Eichler, J. _Haloferax volcanii_ N-glycosylation: delineating the pathway
of dTDP-rhamnose biosynthesis. _Plos One_ 9, e97441 (2014). PubMed PubMed Central Google Scholar * Van der Beek, S. L. et al. Streptococcal dTDP-L-rhamnose biosynthesis enzymes:
functional characterization and lead compound identification. _Mol. Microbiol._ 111, 951–964 (2019). PubMed PubMed Central Google Scholar * Little, K., Tipping, M. J. & Gibbs, K. A.
Swarmer cell development of the bacterium _Proteus mirabilis_ requires the conserved enterobacterial common antigen biosynthesis gene _rffG_. _J. Bacteriol._ 200, e00230–18 (2018). CAS
PubMed PubMed Central Google Scholar * Becker, H. F. et al. Substrate interaction dynamics and oxygen control in the active site of thymidylate synthase ThyX. _Biochem. J._ 459, 37–45
(2014). CAS PubMed Google Scholar * Singh, S. et al. Aldehyde dehydrogenases in cellular responses to oxidative/electrophilic stress. _Free Radic. Biol. Med._ 56, 89–101 (2013). CAS
PubMed Google Scholar * Jackson, B. et al. Update on the aldehyde dehydrogenase gene (ALDH) superfamily. _Hum. Genomics_ 5, 283–303 (2011). CAS PubMed PubMed Central Google Scholar *
Firn, R. D. & Jones, C. G. Natural products - a simple model to explain chemical diversity. _Nat. Prod. Rep._ 20, 382–391 (2003). CAS PubMed Google Scholar * Bleich, R., Watrous, J.
D., Dorrestein, P. C., Bowers, A. A. & Shank, E. A. Thiopeptide antibiotics stimulate biofilm formation in _Bacillus subtilis_. _Proc. Natl Acad. Sci. USA_ 112, 3086–3091 (2015). CAS
PubMed PubMed Central Google Scholar * Romero, D., Traxler, M. F., Lopez, D. & Kolter, R. Antibiotics as signal molecules. _Chem. Rev._ 111, 5492–5505 (2011). CAS PubMed PubMed
Central Google Scholar * Meirelles, L. A. & Newman, D. K. Both toxic and beneficial effects of pyocyanin contribute to the lifecycle of _Pseudomonas aeruginosa_. _Mol. Microbiol._ 110,
995–1010 (2018). CAS PubMed PubMed Central Google Scholar * Will, S. E. et al. The limits to growth - energetic burden of the endogenous antibiotic tropodithietic acid in _Phaeobacter
inhibens_ DSM 17395. _Plos One_ 12, e0177295 (2017). PubMed PubMed Central Google Scholar * Wünsch, D. et al. Global response of _Phaeobacter inhibens_ DSM 17395 to deletion of its 262-kb
chromid encoding antibiotic synthesis. _Microb. Physiol._ 30, 9–24 (2020). PubMed Google Scholar * Zhao, X. H., Yang, X. L. & Hang, H. C. Chemoproteomic analysis of microbiota
metabolite-protein targets and mechanisms. _Biochemistry_ 61, 2822–2834 (2022). CAS PubMed Google Scholar * Kavunja, H. W. et al. Photoactivatable glycolipid probes for identifying
mycolate-protein interactions in live mycobacteria. _J. Am. Chem. Soc._ 142, 7725–7731 (2020). CAS PubMed PubMed Central Google Scholar * Forster, E. R., Yang, X., Tai, A. K., Hang, H.
C. & Shen, A. M. Identification of a bile acid-binding transcription factor in _Clostridioides difficile_ using chemical proteomics. _ACS Chem. Biol._ 17, 3086–3099 (2022). CAS PubMed
PubMed Central Google Scholar * Wilkinson, I. V. L., Bottlinger, M., El Harraoui, Y. & Sieber, S. A. Profiling the heme-binding proteomes of bacteria using chemical proteomics. _Angew.
Chem. Int. Ed. Engl._ 62, e202212111 (2023). CAS PubMed Google Scholar * Keller, L. J., Babin, B. M., Lakemeyer, M. & Bogyo, M. Activity-based protein profiling in bacteria:
applications for identification of therapeutic targets and characterization of microbial communities. _Curr. Opin. Chem. Biol._ 54, 45–53 (2020). CAS PubMed Google Scholar * Wright, M. H.
Chemical proteomics of host-microbe interactions. _Proteomics_ 18, e1700333 (2018). PubMed Google Scholar * Ning, X. J., Wang, X. R., Wu, Y. T., Kang, Q. J. & Bai, L. Q.
Identification and engineering of post-PKS modification bottlenecks for ansamitocin P-3 titer improvement in _Actinosynnema pretiosum_ subsp. _pretiosum_ ATCC 31280. _Biotechnol. J_. 12
https://doi.org/10.1002/biot.201700484 (2017). * Wang, X. R. et al. Efflux identification and engineering for ansamitocin P-3 production in _Actinosynnema pretiosum_. _Appl. Microbiol.
Biotechnol._ 105, 695–706 (2021). CAS PubMed Google Scholar * Pietrocola, F., Galluzzi, L., Bravo-San Pedro, J. M., Madeo, F. & Kroemer, G. Acetyl coenzyme A: a central metabolite and
second messenger. _Cell Metab._ 21, 805–821 (2015). CAS PubMed Google Scholar * Milke, L. & Marienhagen, J. Engineering intracellular malonyl-CoA availability in microbial hosts and
its impact on polyketide and fatty acid synthesis. _Appl. Microbiol. Biotechnol._ 104, 6057–6065 (2020). CAS PubMed PubMed Central Google Scholar * Li, Z. Q. et al. Design and synthesis
of minimalist terminal alkyne-containing diazirine photo-crosslinkers and their incorporation into kinase inhibitors for cell- and tissue-based proteome profiling. _Angew. Chem. Int. Ed._
52, 8551–8556 (2013). CAS Google Scholar * Li, W. C. et al. Development of photoaffinity probe for the discovery of steviol glycosides biosynthesis pathway in _Stevia rebuadiana_ and rapid
substrate screening. _ACS Chem. Biol._ 13, 1944–1949 (2018). CAS PubMed Google Scholar * Li, Y. et al. Dual carbamoylations on the polyketide and glycosyl moiety by Asm21 result in
extended ansamitocin biosynthesis. _Chem. Biol._ 18, 1571–1580 (2011). CAS PubMed Google Scholar * Shi, X. X., Sha, S. S., Liu, L. K., Li, X. & Ma, Y. F. A 96-well microtiter plate
assay for high-throughput screening of _Mycobacterium tuberculosis_ dTDP-D-glucose 4,6-dehydratase inhibitors. _Anal. Biochem._ 498, 53–58 (2016). CAS PubMed Google Scholar * Li, C. Z. et
al. Amino acid catabolism regulates hematopoietic stem cell proteostasis via a GCN2-eIF2α axis. _Cell Stem Cell_ 29, 1119–1134 (2022). CAS PubMed Google Scholar * Sarkar, A. et al.
_Mycobacterium tuberculosis_ thymidylate synthase (ThyX) is a target for plumbagin, a natural product with antimycobacterial activity. _Plos One_ 15, e0228657 (2020). CAS PubMed PubMed
Central Google Scholar * Zhou, Y. Q. et al. Chemical proteomics reveal CD147 as a functional target of pseudolaric acid B in human cancer cells. _ChemComm_ 53, 8671–8674 (2017). CAS
Google Scholar * Jumper, J. et al. Highly accurate protein structure prediction with AlphaFold. _Nature_ 596, 583–589 (2021). CAS PubMed PubMed Central Google Scholar * Schrodinger, L.
L. C., The PyMOL molecular graphics system, version 1.8. (2015). * Ravindranath, P. A., Forli, S., Goodsell, D. S., Olson, A. J. & Sanner, M. F. AutoDockFR: advances in protein-ligand
docking with explicitly specified binding site flexibility. _Plos Comput. Biol._ 11, e1004586 (2015). PubMed PubMed Central Google Scholar * Eberhardt, J., Santos-Martins, D., Tillack, A.
F. & Forli, S. AutoDock Vina 1.2.0: new docking methods, expanded force field, and python bindings. _J. Chem. Inf. Model._ 61, 3891–3898 (2021). CAS PubMed Google Scholar *
Laskowski, R. A. & Swindells, M. B. LigPlot+: multiple ligand-protein interaction diagrams for drug discovery. _J. Chem. Inf. Model._ 51, 2778–2786 (2011). CAS PubMed Google Scholar
Download references ACKNOWLEDGEMENTS We dedicate this paper to the memory of our late colleague, Prof. Youli Xiao. We acknowledge the early efforts by Youli Xiao to synthesize the
photoaffinity probe and conduct the chemoproteomic analysis. We appreciate the helpful comments from Prof. Shenying Li of Shandong University and Prof. Ningyi Zhou from SJTU. We thank the
Core Facility and Technical Service Center for SLSB and the Instrumental Analysis Center in SJTU for data collection. This work was supported by the National Key Research and Development
Program of China (grant Nos. 2021YFC2100600 and 2019YFA0905400), National Natural Science Foundation of China (grant No. 31830104), and Science and Technology Commission of Shanghai
Municipality (grant Nos.19JC1413000 and 19430750600) to L.B. AUTHOR INFORMATION Author notes * These authors jointly supervised this work: Qianjin Kang, Linquan Bai. AUTHORS AND AFFILIATIONS
* State Key Laboratory of Microbial Metabolism, Shanghai-Islamabad-Belgrade Joint Innovation Center on Antibacterial Resistances, School of Life Sciences and Biotechnology, Shanghai Jiao
Tong University, Shanghai, 200240, China Qungang Huang, Xin Zhang, Ziyue Guo, Xinnan Fu, Yilei Zhao, Qianjin Kang & Linquan Bai * Joint International Research Laboratory of Metabolic and
Developmental Sciences, Shanghai Jiao Tong University, Shanghai, 200240, China Qungang Huang, Xin Zhang, Ziyue Guo, Xinnan Fu, Yilei Zhao, Qianjin Kang & Linquan Bai Authors * Qungang
Huang View author publications You can also search for this author inPubMed Google Scholar * Xin Zhang View author publications You can also search for this author inPubMed Google Scholar *
Ziyue Guo View author publications You can also search for this author inPubMed Google Scholar * Xinnan Fu View author publications You can also search for this author inPubMed Google
Scholar * Yilei Zhao View author publications You can also search for this author inPubMed Google Scholar * Qianjin Kang View author publications You can also search for this author inPubMed
Google Scholar * Linquan Bai View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS L.Q.B. and Q.G.H. conceived the project. L.Q.B., Q.J.K., and
Q.G.H. designed the experiments. Q.G.H., X.Z., X.N.F. and Z.Y.G. performed the experiments. Q.G.H., Q.J.K., Y.L.Z. and L.Q.B. analyzed the data. Q.G.H., Q.J.K., Y.L.Z., and L.Q.B. wrote the
manuscript. CORRESPONDING AUTHORS Correspondence to Qianjin Kang or Linquan Bai. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. PEER REVIEW PEER REVIEW
INFORMATION _Communications Biology_ thanks Zhiqiang Cai, Constance B. Bailey, and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling
Editors: [Tobias Goris and Anam Akhtar]. A peer review file is available. ADDITIONAL INFORMATION PUBLISHER’S NOTE Springer Nature remains neutral with regard to jurisdictional claims in
published maps and institutional affiliations. SUPPLEMENTARY INFORMATION PEER REVIEW FILE SUPPLEMENTARY INFORMATION DESCRIPTION OF ADDITIONAL SUPPLEMENTARY FILES SUPPLEMENTARY DATA 1
SUPPLEMENTARY DATA 2 REPORTING SUMMARY RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing,
adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons
licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a
credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted
use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT
THIS ARTICLE CITE THIS ARTICLE Huang, Q., Zhang, X., Guo, Z. _et al._ Biosynthesis of ansamitocin P-3 incurs stress on the producing strain _Actinosynnema pretiosum_ at multiple targets.
_Commun Biol_ 6, 860 (2023). https://doi.org/10.1038/s42003-023-05227-w Download citation * Received: 26 April 2023 * Accepted: 07 August 2023 * Published: 18 August 2023 * DOI:
https://doi.org/10.1038/s42003-023-05227-w SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not
currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative