- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT The Fukushima Daiichi Nuclear Power Plant accident caused the release of large amounts of radioactive material into the environment. Radiation from radionuclides cause DNA lesions,
mainly via oxidation, which adversely affect wild organisms by damaging their germ cells. Here, we investigated the effects of radiation on the reproductive organs of Japanese field mice
(_Apodemus speciosus_) by estimating the dose rate of radiation exposure, the accumulation of DNA lesions, and the expression of DNA repair enzymes. In highly contaminated areas, mouse
testes received a radiation dose rate > 0.1 mGy/d. According to the International Commission on Radiological Protection, there is a very low probability of effects in the reference rat
species at this exposure level. The results of the current study do not definitively conclude that the expression of 8-oxoguanine DNA glycosylase 1 and superoxide dismutase in mouse testes
increase with dose rate and lifetime dose. However, 8-hydroxy-2′-deoxyguanosine accumulation increases in a dose rate- and lifetime dose-dependent manner in mouse testes, but is not observed
in the sperm of the cauda epididymis. These results suggest that, although DNA lesions occurred in male germ cells of Fukushima mice, most were successfully repaired by DNA repair enzymes
at the observed gene expression level. SIMILAR CONTENT BEING VIEWED BY OTHERS DIAZINON INDUCES TESTICULAR DYSFUNCTION AND TESTICULAR CELL DAMAGE THROUGH INCREASED REACTIVE OXYGEN SPECIES
PRODUCTION IN MOUSE Article Open access 21 March 2025 BIOMARKER DOSIMETRY OF ACUTE LOW LEVEL OF THERMAL NEUTRONS AND RADIATION ADAPTIVE RESPONSE EFFECT ON RATS Article Open access 09 August
2024 INVESTIGATING THE IMPACT OF LONG TERM EXPOSURE TO CHEMICAL AGENTS ON THE CHROMOSOMAL RADIOSENSITIVITY USING HUMAN LYMPHOBLASTOID GM1899A CELLS Article Open access 16 June 2021
INTRODUCTION The Fukushima Daiichi Nuclear Power Plant (FDNPP) accident on March 11, 2011, released approximately 7,500 PBq of radioactive materials into the environment1. The Japanese
government designated the areas where the annual cumulative radiation dose estimated from the air dose rate exceeded 50 mSv and the annual accumulated dose was not likely to decline below 20
mSv even after five years as ‘difficult-to-return zones’ in 2012. Although humans evacuated these difficult-to-return zones, wild flora and fauna remained and were exposed to radiation.
Previous studies have attempted to estimate radiation exposure levels in terrestrial organisms in difficult-to-return zones near the FDNPP2,3,4,5,6,7,8,9,10. Garnier-Laplace et al.8 showed
that forest rodents were exposed to radiation at a rate of 3.9 mGy/d during the first 30 d after the accident. Radiation exposure can reduce reproductive success, according to the
International Commission on Radiological Protection (ICRP). Strand et al.9 estimated that terrestrial wild organisms inhabiting areas near the FDNPP were exposed to 100–300 µGy/h (2.4–7.2
mGy/d) for the first three months after the accident. Almost all radiation is emitted by short-lived radioisotopes such as iodine-131 (T1/2 = 8 d)9. For at least three months after the
accident, wild organisms were exposed to just 10–70 µGy/h (0.24–1.68 mGy/d) due to the radioactive decay of short-half-life contaminants9. Thereafter, Cs-134 and Cs-137 became the main
sources of radiation to which wild organisms were exposed in Fukushima. These isotopes have longer half-lives (T1/2 = 2 and 30 years, respectively). The ICRP defines a set of reference
species and recommends dosages for assessing and managing radiation exposure in wild organisms11. According to an ICRP report, mammals are the most sensitive of all organisms to radiation
and show symptoms when exposed to > 0.1 mGy/d. Currently, more data on exposure are available for rodents than for other mammals11. In Japan, a candidate reference rodent species is the
large Japanese field mouse (_Apodemus speciosus_). It is a medium-sized mouse with a long tail, large eyes, and a brown or orange-brown coat12, that dwells primarily on the ground but
occasionally climbs trees12. Therefore, the species may be more exposed to radiation accumulated on the ground than other animals that are arboreal or have a longer distance between the soil
surface and the body. The Japanese field mouse inhabits a wide range of habitats ranging from open fields to secondary lowland forests. Its mean home range is 304–1,853 m2, smaller than
that of other terrestrial wild mammals12,13. This characteristic makes it possible to evaluate the effects of contaminants, such as ionising radiation, in the local environment. The
biological effects of ionising radiation are well-known and can be categorised into two types. The first category includes deterministic effects that appear when the radiation dose exceeds a
threshold; responses include ulcerations, permanent infertility, and death. The other category includes stochastic effects such as carcinogenesis. These effects occur in a dose-dependent
manner, and resulting traits are inherited by the progeny of affected parents in the form of DNA lesions. Stochastic effects occur either directly or indirectly due to radiation exposure.
Recent studies on the absorbed radiation dose rate in wild rodents living in radiation-contaminated areas have shown that the contribution of external exposure, which is composed of gamma
rays, is greater than that of the internal dose rate4,10. Gamma rays, a type of low-linear-energy transfer radiation, produce more indirect effects than direct effects14. For example,
radiation generates reactive oxygen species (ROS) such as superoxide radicals (O2−), hydroxyl radicals (OH·), and hydrogen peroxide (H2O2) from water radiolysis. OH· is highly reactive and
can produce several DNA modifications15,16,17. The most abundant and studied oxidative DNA base modification is 8-hydroxy-2′-deoxyguanosine (8-OHdG), which is produced by the reaction of OH·
with guanine16,18. 8-OHdG pairs with adenosine and cytosine during DNA replication and causes a G: C to T: A transversion mutation19. If such radiation-induced mutations occur in the germ
cells of organisms inhabiting areas exposed to high radiation levels, the accumulated unexpected mutations can be inherited by their progeny, and the likelihood of hereditary diseases
increases. However, organisms possess mechanisms for eliminating ROS and repairing DNA that counteract mutagenesis. Antioxidant enzymes such as superoxide dismutase (SOD), glutathione
peroxidase, and catalase eliminate radiation-induced ROS. When DNA is damaged by radiation, DNA glycosylases that participate in base excision, such as 8-oxoguanine DNA glycosylase (_OGG1_),
repair the DNA damage by hydrolysing oxidised deoxynucleotides (dNTPs). _OGG1_ hydrolyses the N-glycosyl bond, releases 8-OHdG as a free base, and cleaves the sugar-phosphate
backbone17,20,21. In mice, the highest expression level of _OGG1_ mRNA is observed in the testes, implying that _OGG1_ plays an important role in minimising germline mutations21,22. _OGG1_
mRNA expression is elevated by oxidative stressors23,24,25. This study determined whether radiation-induced oxidation following the FDNPP accident affected the reproductive organs of large
Japanese field mice. We compared the effects of radiation exposure on the testes of field mice captured near the highly contaminated Fukushima area, designated by the Japanese government as
a difficult-to-return zone, with that in mice captured in other (uncontaminated) prefectures. Testicular _SOD1_ expression, 8-OHdG accumulation, and _OGG1_ transcription were compared among
field mice captured from areas with different radiation levels. RESULTS DAILY DOSE RATE AND TOTAL LIFETIME DOSE IN MOUSE TESTIS Cs-134 and Cs-137 concentrations were higher in the bodies of
mice from Fukushima than in those from Toyama and Aomori throughout the experimental period (Table 1). Radioactive Cs concentrations were transformed into internal doses in the mouse testes
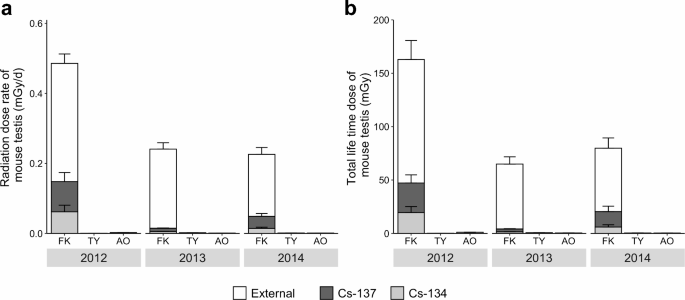
using the Electron Gamma Shower (EGS5) Monte Carlo simulation program. In 2012, at Fukushima, the average daily internal Cs-134 and Cs-137 dose rates and the external dose rate from
radioactive substances in the environment in testis were 0.061 ± 0.019, 0.086 ± 0.026, and 0.338 ± 0.027 mGy/d, respectively (Fig. 1a). The average daily internal dose rates of Cs-134 and
Cs-137 rapidly decreased to 0.005 ± 0.000 and 0.010 ± 0.001 mGy/d in 2013 and 0.014 ± 0.003 and 0.034 ± 0.008 mGy/d in 2014, respectively. A gradual decrease in the average external dose
rate (0.226 ± 0.018 mGy/d in 2013 and 0.177 ± 0.020 mGy/d in 2014) was observed. The ratios of the internal dose rate to the total dose rate in mouse testes were 30.4% in 2012, 6.1% in 2013,
and 21.5% in 2014 (Fig. 1a), suggesting that radiation exposure in mouse testes was mainly external rather than internal. In contrast to Fukushima, the total dose rate at Toyama and Aomori
was < 0.005 mGy/d throughout the study period. Internal dose rates were < 10% of the total dose rate (Toyama: 1.5% in 2012, 0.1% in 2013, and 0.4% in 2014; Aomori: 7.2% in 2012, 1.6%
in 2013, and 9.0% in 2014). The lifetime dose to which each mouse testis was exposed was calculated using the animal’s age derived from the degree of tooth wear (Fig. 1b). The average total
lifetime dose of mouse testis in Fukushima was 184.7 ± 26.4 mGy in 2012, 65.0 ± 6.7 mGy in 2013, and 80.0 ± 9.6 mGy in 2014 (Fig. 1b). In the control groups, the 2012 Aomori sample showed a
total lifetime dose of 1.1 ± 0.1 mGy in the testis, but other samples had values below 1 mGy (Fig. 1b). Similar values and tendencies were observed when calculating whole-body exposure
(Supplementary Fig. S1). EVALUATION OF OXIDATIVE STRESS IN MOUSE TESTES For Fukushima samples, we first measured _Sod1_ expression. _Sod1_ encodes an enzyme that removes radiation-generated
free radicals. _Sod1_ transcription levels in the testes of mice captured in Fukushima in 2012 were higher than those in mice from control sites. A significant difference in _Sod1_
transcription was observed between Fukushima and Aomori (Steel–Dwass; Fukushima (FK) vs. Toyama (TY): _W_ = −3.259, _p_ = 0.051; FK vs. Aomori (AO): _W_ = −4.472, _p_ < 0.001; TY vs. AO:
_W_ = −1.039, _p_ = 0.794; Fig. 2a). In 2013 and 2014, testicular _Sod1_ expression in Fukushima mice decreased, but the values were not significantly different from those in Toyama and
Aomori mice (Steel–Dwass; 2013, FK vs. TY: _W_ = 0.000, _p_ = 1.000, FK vs. AO: _W_ = −2.877, _p_ = 0.103, TY vs. AO: _W_ = −2.210, _p_ = 0.265; 2014, FK vs. TY: _W_ = 1.244, _p_ = 0.670, FK
vs. AO: _W_ = −0.387, _p_ = 0.979, TY vs. AO: _W_ = −0.956, _p_ = 0.815; Fig. 2a). We also investigated the accumulation of 8-OHdG in the testes and cauda epididymis to elucidate the extent
of free radical DNA oxidation (Fig. 2b). The percentage of 8-OHdG-positive cells in the testicular seminiferous tubules of mice from Fukushima in 2012 was significantly higher than that of
mice from Toyama and Aomori (Steel–Dwass; FK vs. TY: _W_ = −4.472, _p_ < 0.001; FK vs. AO: _W_ = −5.020, _p_ = 0.000; TY vs. AO: _W_ = 0.1148, _p_ = 1.000; Fig. 2c). Overall, the
proportion of accumulated 8-OHdG-positive cells decreased from 2012 to 2013; however, the proportion of accumulated 8-OHdG-positive cells in Fukushima mice remained significantly higher than
that in mice at other sites (Steel–Dwass; FK vs. TY: _W_ = −4.855, _p_ < 0.001; FK vs. AO: _W_ = −6.121, _p_ = 0.000; TY vs. AO: _W_ = −2.242, _p_ = 0.261; Fig. 2c). In 2014, no maximum
proportion of accumulated 8-OHdG-positive cells was found in mice captured in Fukushima, nor was there a significant difference in the proportion of 8-OHdG-positive cells between Fukushima
and Aomori mice (Steel–Dwass; FK vs. TY: _W_ = −4.768, _p_ = 0.000; FK vs. AO: _W_ = −0.817, _p_ = 0.861; TY vs. AO: _W_ = 2.191, _p_ = 0.313; Fig. 2c). When the positive cell rate was
tabulated for each spermatogenesis stage using the data from five individuals with a particularly high 8-OHdG-positive cell rate, a high percentage of positive cells was found in stages 9–14
(Supplementary Fig. S2). In contrast, no positive cells were found in the cauda epididymis of mice captured in Fukushima in 2012 or 2013. In 2012, the transcript level of _Ogg1_ was five
times higher in Fukushima mouse testes than in those of mice from other sites (Steel–Dwass; FK vs. TY: _W_ = −4.116, _p_ = 0.003; FK vs. AO: _W_ = −4.472, _p_ < 0.001; TY vs. AO: _W_ =
−2.079, _p_ = 0.343; Fig. 2d). In contrast, there were no significant differences in _Ogg1_ transcript levels between the sites in 2013 and 2014. In 2014, however, the expression of this
gene was slightly higher in the testes of mice from Fukushima than in those from other sites (Steel–Dwass; 2013, FK vs. TY: _W_ = 0.074, _p_ = 1.000, FK vs. AO: _W_ = 0.662, _p_ = 0.897, TY
vs. AO: _W_ = 0.375, _p_ = 0.966; 2014, FK vs. TY: _W_ = −0.560, _p_ = 0.936, FK vs. AO: _W_ = −1.291, _p_ = 0.673, TY vs. AO: _W_ = −1.673, _p_ = 0.514; Fig. 2d). RELATIONSHIP BETWEEN
OXIDATIVE STRESS MARKERS, RADIATION DOSE RATE, AND LIFETIME DOSE IN TESTIS The contribution of radiation to oxidative stress marker induction was examined by constructing linear
mixed-effects models that included thresholds (Table 2). Akaike’s information criterion with correction (AICc) scores were determined without _Sod1_ or the lifetime dose in the mouse testis
and body (Supplementary Methods and Supplementary Fig. S3). Subsequently, a model with a threshold was reconstructed. If the threshold value is in a range, the model was constructed using
the median value. The coefficient values indicated a significant relationship between the daily dose rate in the testis and _Sod1_ mRNA expression, the ratio of 8-OHdG-positive cells, and
_Ogg1_ mRNA expression (Table 2; Fig. 3a), and between the lifetime dose in the testis and the ratio of 8-OHdG-positive cells and _Ogg1_ mRNA expression (Table 2; Fig. 3b). For models of
_Sod1_ and _Ogg1_, the assumption of normality of residuals was met (Shapiro–Wilk normality test; Table 2). A similar tendency was observed when calculating the whole-body exposure
(Supplementary Fig. S4 and Supplementary Table S1). DISCUSSION This study serves as an addendum to our previous work on the effects of radiation on wild large Japanese field mice26. In the
present study, we also included data from 2012, a year with higher radiation levels than 2013 and 2014, and calculated specific exposure levels (daily dose rate and total lifetime dose). The
previous report only noted the ambient dose rates of trapping sites and caesium accumulation levels within the mouse bodies. The specific daily dose rate and the total lifetime dose were
not calculated. To accurately understand the effects of radiation on biological processes, it is essential to calculate both the specific daily dose rate and the total lifetime dose. The
previous study on mice captured in difficult-to-return zones in 2013 and 2014 reported that the apoptotic cell frequencies and the frequencies of morphologically abnormal sperm when compared
with non-contaminated sites showed no significant statistical difference. In this study, we used oxidative stress markers to evaluate DNA damage caused by radiation. The testes of large
Japanese field mice collected from Fukushima were exposed to higher radiation dose rates than those of large Japanese field mice collected from the control sites. The external dose rate
accounted for approximately 70% of the total dose rate. These results suggest that the Japanese field mice in Fukushima were mainly exposed to radiation from their habitats. Our results
showed that internal exposure (dose rate and total lifetime dose, Fig. 1a, b) to Cs-134 and Cs-137 was higher in 2014 than 2013. This observed trend may be attributed to a transient
elevation in the Cs concentration in the soil resulting from the deposition of falling leaves and rainwater runoff at the capture site. This phenomenon may affect the Cs concentrations in
the food of mice, increasing their internal exposure levels. In 2012, _Sod1_ transcription, 8-OHdG accumulation, and _Ogg1_ transcription in the testes of Fukushima mice were significantly
greater than those in the testes of mice from control sites; however, the differences were no longer clear in 2013 and 2014 (Fig. 2a, c, d), presumably because of the small sample sizes in
each study area and annual decreases in the total radiation dose rate. To clarify the general trends in the relationships between the radiation dose rate and levels of oxidative stress
markers, we constructed linear mixed-effects models incorporating year and site as random effects. The same model was also constructed to determine the relationship between the lifetime dose
and oxidative stress markers. The models, including the threshold, were considered the best for all stress markers according to the AICc information criterion (Supplementary Fig. S3,
Supplementary Table S2). For the relationship between markers _Sod1_ and _Ogg1_ and the daily dose rate in mouse testes, the _Sod1_ and _Ogg1_ models were successfully constructed but showed
no significant or negative intercepts and initial slopes (Table 2). Additionally, there was a threshold, but it was close to zero, due to the distribution of the data and lack of biological
significance (Fig. 3a). Therefore, care must be taken when interpreting the meaning of the thresholds. When models were analysed by year, no significant _Sod1_ and _Ogg1_ models were
established (Supplementary Data S1). One reason why a significant model could be constructed with combined data from 3 years but not with data from each year may be the annual environmental
fluctuations during the trapping period (i.e., the breeding season). For example, temperature, rainfall, and food quality during the breeding season varied each year, which may have affected
gene expression levels at the time of capture. By combining data from three years, the effect of these annual variations on the results can be mitigated. In contrast, when analysing data
from each year separately, these variations may have a substantial impact, making it more difficult to obtain significant results. Considering that the maximum reported lifespan for the
Japanese field mouse is 26 months12, combining the data from 3 years in our analysis averaged out these environmental factors. However, considering the discrepancies between the results of
the combined three-year analysis and the yearly analyses, we could not definitively infer that the expression levels of _Sod1_ and _Ogg1_ increase in a dose-rate-dependent manner. For
8-OHdG, a threshold was observed at 0.18 to 0.19 mGy/d for the daily dose rate in mouse testis (Supplementary Fig. S3 and Supplementary Table S2). The ratio of 8-OHdG-positive cells
increased around this threshold, with the response being initially strong and slowing after the threshold (Fig. 3a). When 8-OHdG data were analysed for each year, models were constructed in
2012, 2013, and 2014 and a significant relationship between the daily dose rate and the ratio of 8-OHdG-positive cells in 2012 and 2013 was confirmed (Supplementary Data S1). After pooling
three years of data, it was confirmed that significant models could also be developed as mentioned above. Therefore, the proportion of 8-OHdG-positive cells increases in a
dose-rate-dependent manner. During the study period in Fukushima, the average internal and external radiation dose rates ranged from 0.1 to 1.0 mGy/d. According to the ICRP11,27, such a
level of radiation induces certain effects in reference rats with a very low probability. The result suggested that one of the effects of radiation in large Japanese field mice in Fukushima
could be the accumulation of 8-OHdG. To determine the relationship between markers and the total lifetime dose in mouse testes, a linear mixed-effect model for _Sod1_, _Ogg1_, and the
proportion of 8-OHdG-positive cells was attempted. The _Sod1_ model was not constructed (Table 2). On the other hand, significant _Ogg1_ models were successfully constructed (Table 2). When
models were analysed by year, no significant models relevant to _Sod1_ and _Ogg1_ were established (Supplementary Data S1). These results suggested that _Sod1_ expression does not increase
with lifetime dose. The transcriptional regulation of _Sod1_ is highly controlled, and mRNA levels of the gene increase in response to various stressors such as chemicals, heat shock, and
irradiation28. Increased _Sod1_ transcription in response to elevated ROS levels has also been reported29,30. As oxidative stress markers respond quickly to stimuli, the dose rate per unit
time may be a better fit than the total integrated dose. With regard to _Ogg1_, annual environmental fluctuations affected the construction of models, similar to their effect on the
construction of _Sod1_ and _Ogg1_ models in relation to daily dose rate. When analyzing data from each year separately, these fluctuations may have a substantial impact, making it more
difficult to obtain significant results. In contrast, analyzing combined data from three years averaged out these environmental factors. However, considering the discrepancies between the
results of the combined three-year analysis and the yearly analyses, we believe that these results do not clearly indicate that the expression levels of _Ogg1_ increase in a total lifetime
dose-dependent manner. Although it is unclear whether _Ogg1_ increases in a dose-dependent manner, as will be shown later, it is likely that DNA repair occurs at the observed _Ogg1_
expression levels. Analysis of 8-OHdG also showed a threshold of 94–105 mGy for the lifetime dose in mouse testes. The ratio of 8-OHdG-positive cells increased around this threshold, with
the response being initially strong and slowing after the threshold (Fig. 3b). When 8-OHdG data were analysed for each year, models were constructed in 2012, 2013, and 2014, and a
significant relationship between the lifetime dose and the ration of 8-OHdG-positive cells in 2012 was confirmed (Supplementary Data S1). After pooling three years of data, it was confirmed
that significant models could also be developed as mentioned above. Therefore, we concluded that the ration of 8-OHdG-positive cells is considered to increase in a lifetime dose-dependent
manner. In addition, the results of X-ray irradiation experiments at a dose of 100 mGy using captive-bred Japanese field mice showed 8-OHdG accumulation in the testes of mice (Supplementary
Note and Supplementary Data S1, S2 and S3). This result suggested that the phenomenon of a high level of 8-OHdG accumulation observed in the testes of mice from Fukushima was due to the
radiation effect of radioactive materials in the environment. A high percentage of 8-OHdG-positive cells was found in spermatogenesis stages 9–14 in the testis, which suggests that stage
9–14 spermatocytes are highly sensitive to 8-OHdG. The accumulation of 8-OHdG leads to an increase in DNA mutations and carcinogenesis31,32,33,34. Previous studies have shown that
_Ogg1_-deficient mice exhibit elevated hepatic 8-OHdG accumulation and DNA mutation frequencies31,32. Mice lacking mutT homologue 1 (_Mth1_), _Ogg1_, and mutY homologue (_Mutyh_), which
encode base excision repair enzymes, exhibit an increased number of base substitution mutations from G to T in the germline33 and a higher lung tumor incidence34. Additional supportive
information has shown that this may increase mutation frequency in wild rodents distributed in areas contaminated with radioactive materials. Chromosomal aberrations have also been observed
in wild rodents in contaminated areas. For example, small Japanese field mice (_Apodemus argenteus_) and house mice (_Mus musculus_) collected from Fukushima Prefecture were exposed to
higher radiation levels (approximately 1–3 mGy/d) than those observed in the present study (maximum 0.91 mGy/d). The frequency of chromosomal aberrations in splenic lymphocytes tended to
increase with increasing dose rate4. Based on the above-reported data, large Japanese field mice inhabiting areas with higher radiation exposure than those in the present study may exhibit
DNA mutations. In contrast with the testicular results, 8-OHdG accumulation was not detected in the cauda epididymis of the mice captured in Fukushima. The dose rate and lifetime dose
dependency of _Ogg1_ expression in mouse testes were not clear; however, the base excision repair system in large Japanese field mice functioned effectively even in highly contaminated areas
of Fukushima, with most 8-OHdG likely repaired at the observed _Ogg1_ expression levels. Therefore, DNA mutations in sperm cells were effectively minimised by the DNA repair system in large
Japanese field mice in the study area. To confirm this, it is necessary to compare the genomic information of sperm cells in the cauda epididymis with that of somatic cells to confirm that
DNA repair worked effectively. Moreover, extending our research focus, members of our team are currently investigating mutations in proteins related to DNA damage repair pathways. This
research aims to understand how repair mechanisms for DNA and double-strand breaks operate under conditions of chronic radiation exposure. By exploring mutations in DNA repair proteins, we
hope to elucidate how these proteins adjust under radiation stress and whether they are altered in a way that affects their function. According to its ecological life history, the Japanese
field mouse has two reproductive seasons: early spring and late summer35,36. The reported maximum lifespans in the wild are approximately 15 months in Hokkaido and 26 in Honshu, Japan.
Considering the information and the estimated age in days, the field mice captured in 2012 represented the first generation exposed to the nuclear accident, the second generation (born in
the summer of 2011), and the third generation (born in the spring of 2012). Similarly, field mice captured in 2013 were categorised as individuals from the third, fourth (born in the summer
of 2012), and fifth generations (born in the spring of 2013). Finally, the field mice captured in 2014 were classified as individuals of the fifth, sixth (born in the summer of 2013), and
seventh generations (born in the spring of 2014). In this study, oxidative stress markers in the mice captured in 2012 showed a clear reaction to radiation. As the dose rate declined, from
March 2011 to August 2012, at least three generations of male mice showed signs of oxidative stress in their reproductive systems. Especially in the first generation, immediately after the
accident, there may have been severe reproductive suppression, according to the radiation dose estimated from soil samples by Garnier-Laplace et al.7. Our results indicate that the effects
of oxidative stress have decreased since 2013. However, a previous study reported enhanced spermatogenesis in mice captured in 201437. The population of field mice inhabiting Akogi (external
dose rate from radiocesium: 407–447 µGy/d), close to the study site of the present study, experienced an increase in the number of spermatogenic and proliferating
cell-nuclear-antigen-positive cells per seminiferous tubule when compared with that observed in areas with a lower external dose rate—Tanashio (21 μGy/d) and Murohara (304~365 μGy/d). Thus,
although oxidative stress levels tend to decrease, other markers, such as spermatogenesis and DNA mutations, can be used to assess the effects of radiation. Furthermore, the potential for
radiation to cause genetic instability and its implications for future generations are of considerable concern. Genetic instability may indirectly cause transgenerational effects38.
Additionally, oxidative stress is implicated in the development of genomic instability39. Therefore, although there are various limitations to research on wild animals, further research
should be conducted on wild animals in combination with laboratory experiments to understand the impact of the Fukushima accident on wildlife, including how successive generations of field
mice may carry genetic alterations initiated by their ancestors’ exposure to radiation forward. In conclusion, this study showed that the testes of mice captured in a radiation-contaminated
area of Fukushima were exposed to approximately 0.1–1.0 mGy/d, which, according to the ICRP, has a very low probability of affecting reproduction. Regarding the expression levels of _Sod1_
and _Ogg1_ in mouse testes, the study findings do not clearly conclude that they increase with increasing dose rate and lifetime dose. Significant accumulation of 8-OHdG in the testes of
field mice inhabiting difficult-to-return zones in Fukushima was observed in 2012 and 2013; however, no 8-OHdG-positive cells were observed in the cauda epididymis. Thus, it is likely that
DNA repair was effectively performed by enzymes in the testes, such as _Ogg1_. Although the accumulation of 8-OHdG increases DNA mutations, DNA mutations in sperm cells can be effectively
minimised by the DNA repair system. To confirm this, it is necessary to compare the genomic information of sperm with that of somatic cells from mice captured in a radiation-contaminated
area of Fukushima, which will confirm whether the mutation rate in the germ cells of the field mice distributed in the difficult-to-return zone was changed by the released radioactive
material and whether a trans-generational effect exists. METHODS STUDY AREA Target organs of large Japanese field mice were sampled during each breeding season (July to September) from 2012
to 2014 (Supplementary Data S4) at three forest sites in Japan. If the number of individuals captured during the study period was small, animals captured in the spring breeding season
(March–April) were used to satisfy the sample size for statistical analysis. One of the sites was a radiation-contaminated area in Fukushima Prefecture (37°35ʹN, 140°44ʹE). The Japanese
government designated this region as a difficult-to-return zone since it was affected by the nuclear power plant accident. Two other sites, one in Aomori Prefecture (40°35ʹN, 140°57ʹE) and
the other in Toyama Prefecture (36°35ʹN, 137°24ʹE), were used as controls. The gamma radiation air dose rates at ground level were measured simultaneously with mouse sampling using a
portable environmental gamma survey meter (NHE20CY3-131By-S; Fuji Electric Co., Ltd., Tokyo, Japan). Air dose rate data were used to estimate the external daily radiation dose rate, as
described below. ANIMALS The mice were captured using Sherman-type live traps baited with sunflower seeds. At each study site, 50–80 traps were set at 10 m intervals within a 2 km radius
from each coordinate. Captures were conducted once or five times per year. Each sampling period lasted 2–3 d. The traps were checked every morning, and captured mice were immediately
transported to the laboratory. Sexually mature males weighing > 30 g40, having paired testes weighing > 0.95 g, and being reproductively active36 were used in the experiments described
below. Mice were euthanised by CO2 asphyxiation and weighed. The testes and cauda epididymis were removed from each mouse and weighed. Half of one testis from each animal was fixed in Bouin
solution at room temperature, and the other half was stored in QIAzol Lysis Reagent (QIAGEN, Hilden, Germany) at −20 °C. After removing the head and internal organs, the body was stored at
− 20 °C. The heads were removed for age determination. The ages of the mice in months were estimated based on molar wear and presented as median age in days (Supplementary Table S3). Mouse
bodies without internal organs or heads were used to measure the mouse body Cs concentration. Tissue was minced using a food processor, packed in a 100 mL polystyrene container, and sample
weight and volume were measured. The Cs-134 and Cs-137 activities of the samples were measured using high-purity germanium (HpGe) detectors (GMX45P4-76, ORTEC, TN or GCW7023, Canberra
Industries Inc., Marion, CT, USA). Gamma Studio software (SEIKO EG&G Co., Ltd., Tokyo, Japan) and Spectrum Explorer software (Canberra) were used to analyse the gamma-ray spectra. A
standard radiation source (MX033U8PP, Japan Radioisotope Association, Tokyo, Japan) was used for the calibration. The radioactivity levels of Cs-134 and Cs-137 were corrected for radioactive
decay to obtain expected values at the time of sampling. Values below the detection limit were considered 0 Bq/kg. The Cs concentrations in the mouse bodies were used to estimate the
internal daily radiation dose rate. All rodent capture experiments were approved by the National Institute for Environmental Studies for the analysis and experimentation of environmental
samples contaminated with radioactive materials (Permission No. H24-10-10) and performed according to the guidelines for the use of wild mammals of the Mammal Society of Japan41 and the
Guidelines for the Care and Use of Laboratory Animals of the National Institute for Environmental Studies. The study was carried out in compliance with the ARRIVE guidelines. CALCULATION OF
DAILY RADIATION DOSE RATES IN TESTES The daily radiation dose rate in the testicles (mGy/d) was expressed as the sum of the daily external and internal radiation dose rates. The external
daily radiation dose rate for the whole body was calculated based on the radiation air dose measured using a portable environmental gamma survey meter (NHE20CY3-131By-S; Fuji Electric Co.,
Ltd., Tokyo, Japan). The ambient dose equivalent rate was used to calculate the daily external dose rate using the following formula with the radiation converting factor of γ-ray 1.042:
Daily external dose rate (mGy/d) = (Ambient dose equivalent rate of trapping site [µSv/h]/1.2) × (24 h/1) × (radiation weighting factor of γ-ray/1,000). The air dose rates were assessed at
arbitrarily chosen locations at each site in 2012. Air dose rates were measured at five random locations at intervals of at least 10 m in 2013 and 2014; average values were used. Due to
missing data for Toyama in 2014, the average values measured in 2012 and 2013 were substituted. We confirmed that the obtained results did not differ, and the data for Toyama in 2014 were
hypothesised to be both the maximum value from 2013 and the minimum value from 2012. The dose rate of beta rays from the ground was not included because the maximum range of beta rays from
Cs-134 and Cs-137 in water is < 2 mm43 and the testes were covered with fur (2.8 mm)44 and skin (0.8 mm)45 and protected by the peritoneum and tunica albuginea; therefore, the
contribution of beta rays was considered negligible. Internal daily radiation dose rates in the testicles were estimated using the EGS5 program46 with mouse body Cs concentration data. The
energy distribution of the beta particles generated from Cs-134 and Cs-137 was calculated according to the Fermi theory47 using experimental data48. The shape of the field mouse testes was
regarded as ellipsoid. The minor and major axes were calculated from the relational formula with testis weight (Ishiniwa et al., unpublished data). The composition of the testes was recorded
as TISSUE and SOFT (ICRP; soft tissue), while the tissues surrounding the testes were recorded as ADIPOSE TISSUE (ICRP; adipose tissue). The composition data were obtained from the NIST
website (http://physics.nist.gov/cgi-bin/Star/compos.pl?matno=261 and http://physics.nist.gov/cgi-bin/Star/compos.pl?refer=ap&matno=103). Cs was assumed to be uniformly distributed
throughout the body of the mice. The specific radioactivities of Cs-134 and Cs-137 were determined based on Ge semiconductor detector measurements. In the model, it was assumed that beta
rays were emitted in all directions from the Cs-134 and Cs-137 present. The probability of an interaction between the beta particles and tissue atoms was determined using the EGS5 program.
The radiation dose rate of the testes was calculated from the simulated beta-particle energy deposition within a fixed period. MeV (cm2 g− 1 s− 1) was converted to Gy/h (J kg− 1 h− 1).
CALCULATION OF LIFETIME RADIATION DOSE IN TESTES The lifetime radiation dose (mGy) in the testicles of individual mice was defined as the sum of the lifetime external and internal radiation
doses. The lifetime external radiation dose for each mouse was calculated as the daily external dose rate multiplied by the estimated age in days. The total lifetime internal radiation dose
for each mouse was calculated by multiplying the daily internal dose rate with the estimated age in days. IMMUNOHISTOCHEMISTRY STAINING FOR TESTICULAR 8-OHDG Testes were fixed in Bouin’s
solution and processed for paraffin embedding. Sections were cut at a thickness of 5 μm, dewaxed, washed with phosphate-buffered saline (PBS) for 5 min, and boiled in 10 mM citric acid (pH
6.0) for 5 min in an autoclave. The sections were then rinsed three times with PBS for 2 min. After 30 min of incubation at room temperature with goat serum albumin (3%), the sections were
covered with the primary antibody, mouse anti-8-OHdG monoclonal antibody (1:20, Japan Institute for the Control of Aging, Shizuoka, Japan), and incubated overnight at 4 °C. The sections were
then washed thrice for 2 min with PBS and covered with the secondary antibody, Histofine Simple Stain Rat MAX PO (MULTI; NICHIREI BIOSCIENCES Inc., Tokyo, Japan), for 20 min at room
temperature. The sections were stained with diaminobenzidine (DAB; Histofine DAB Kit; NICHIREI BIOSCIENCES, Inc.). Haematoxylin was used as a counterstain, and the sections were observed
using a NIKON Bioresearch ECLIPSE 80i Microscope. A negative control section was prepared using the same procedure described above, without the primary mouse anti-8-OHdG monoclonal antibody
(Fig. 2b). To calculate the ratio of 8-OHdG-positive cells in the testes, 100 seminiferous tubules were selected from tissue sections of each mouse. The seminiferous tubes were selected such
that the three stages reported by Oakberg49 and Perey et al.50, were represented by one-third each; between 31 and 34 seminiferous tubes were selected from each of the three stages. We then
counted all the cells in the selected seminiferous tubules, classifying them as either 8-OHdG-positive or -negative. The percentage of 8-OHdG-positive cells was calculated by dividing the
number of 8-OHdG-positive cells in selected seminiferous tubules by the total number of cells in the selected seminiferous tubules. For mice with a high percentage of 8-OHdG-positive cells,
data showing the spermatogenesis stages36 with a high percentage of 8-OHdG-positive cells are presented in Supplementary Figure S2. In addition, immunohistochemistry staining for 8-OHdG was
also performed on the cauda epididymis of mice captured in Fukushima in 2012 (_n_ = 11) and 2013 (_n_ = 20). TRANSCRIPT LEVELS OF _SOD1_, _OGG1_, AND _Β-ACTIN_ DETERMINED USING QUANTITATIVE
REAL-TIME PCR Total testicular RNA was isolated using the EZ1 RNA Universal Tissue Kit (QIAGEN). Single-stranded cDNA was synthesised from total RNA (1 µg) using PrimeScript™ Master Mix
(TaKaRa Bio Inc., Shiga, Japan). The transcript levels of _Sod1_, _Ogg1_, and _β-actin_ mRNA were measured by quantitative real-time PCR performed using KOD SYBR® qPCR Mix (TOYOBO CO., LTD.,
Okasa, Japan) and a Light Cycler® 480 (Roche Diagnostics, Basel, Schweiz). The _Sod1_ and _Ogg1_ primers were as follows: As-SOD1_forward, 5′-TCCAT TGAAG ATCGA GTGAT CTC-3′;
As-SOD1_reverse, 5′-TCACA CCACA AGCCA AGC-3′; As-OGG1_forward, 5′-GATTG CCCAT CGTGA CTAC-3′; and As-OGG1_reverse, 5′-CACTG AACAG CACTG CTTG-3′. The _β-actin_ primers51 were as follows:
As-ACTB_forward, 5′-CAGGA TGCAG AAGGA GATTA C-3′ and As-ACTB_reverse, 5′-ACCGA TCCAC ACAGA GT-3′. Real-time PCR amplifications of _Sod1_, _Ogg1_, and _β-actin_ were performed with forty
cycles of denaturing (95 °C, 10 s), annealing (58 °C, 10 s), and extension (68 °C, 30 s) using a single fluorescence measure following initial denaturation (98 °C, 2 min). After the
amplification and quantification cycles, a melting curve analysis (65–95 °C with a heating rate of 0.1 °C/s and continuous measurement of fluorescence) was conducted to validate the specific
PCR products. A standard curve was constructed by plotting the cross point (Cp) against the continuous dilution of a particular sample. The PCR efficiency (E) of one cycle was calculated as
E = 10−1/S (where S is the slope of the straight line between Cp and dilution). The relative concentration was calculated using the following equation: Concentration (SOD1 or OGG1) = E(SOD1
or OGG1)Cp(SOD1 or OGG1)/E(actb)Cp(actb). STATISTICAL ANALYSIS To compare values of oxidative stress markers among study sites each year, we conducted a Steel–Dwass test in R using the
‘NSM3’ software package52. To assess the relationship between the radiation exposure daily dose rate and the degree of oxidative stress response, we used a linear mixed-effect model with the
‘lmerTest’53, ‘lme4’54, and ‘glmmTMB’ software packages55. The same model was used to assess the relationship between the lifetime dose of radiation and oxidative stress response. The
explanatory variables were the total (external + internal) mouse testis radiation, body radiation dose rate, or lifetime dose, and year and site were included as random effects to account
for confounding effects because the environment at each study site could have varied greatly among the three sites or from year to year. We constructed models for the response variables of
_Sod1_ expression, 8-OHdG-positive cell proportion, and _Ogg1_ expression. The linear regression model assumes the normality of error terms56. The sample residuals are useful for assessing
normality instead of error terms57. The normality of the residuals was confirmed using the Shapiro–Wilk test, which is the most powerful normality test57. For the 8-OHdG-positive cell data,
models were constructed using ‘glmmTMB’, which can include the offset. The existence of ‘thresholds’ in the linear regression models was verified using AICc (Supplementary Methods). An _α_
value of 0.05 was used to indicate statistical significance for all tests. All analyses were conducted using R ver. 3.6.258. The above statistical analyses were conducted not only on the
combined data from 2012, 2013, and 2014 but also on the individual data from each year. DATA AVAILABILITY All data used in this study are included in this manuscript and the Supplementary
Materials. REFERENCES * United Nations Scientific Committee on the Effect of Atomic Radiation. Radionuclide releases in Sources, effects and risks of ionizing radiation: UNSCEAR 2013 report
(ed. UNSCEAR) 110–128. United Nations Publication, (2013). * Yamashiro, H. et al. Effects of radioactive caesium on bull testes after the Fukushima nuclear plant accident. _Sci. Rep._ 3,
2850 (2013). Article PubMed PubMed Central Google Scholar * Murase, K., Murase, J., Horie, R. & Endo, K. Effects of the Fukushima Daiichi nuclear accident on goshawk reproduction.
_Sci. Rep._ 5, 9405 (2015). Article CAS PubMed PubMed Central Google Scholar * Kubota, Y. et al. Chromosomal aberrations in wild mice captured in areas differentially contaminated by
the Fukushima Dai-Ichi nuclear power plant accident. _Environ. Sci. Technol._ 49, 10074–10083 (2015). Article ADS CAS PubMed Google Scholar * Fuma, S. et al. Dose rate estimation of the
Tohoku hynobiid salamander, _Hynobius lichenatus_, in Fukushima. _J. Environ. Radioact_. 143, 123–134 (2015). Article CAS PubMed Google Scholar * Bonisoli-Alquati, A. et al. Abundance
and genetic damage of barn swallows from Fukushima. _Sci. Rep._ 5, 9432 (2015). Article CAS PubMed PubMed Central Google Scholar * Garnier-Laplace, J. et al. Radiological dose
reconstruction for birds reconciles outcomes of Fukushima with knowledge of dose-effect relationships. _Sci. Rep._ 5, 16594 (2015). Article ADS CAS PubMed PubMed Central Google Scholar
* Garnier-Laplace, J., Beaugelin-Seiller, K. & Hinton, T. G. Fukushima wildlife dose reconstruction signals ecological consequences. _Environ. Sci. Technol._ 45, 5077–5078 (2011).
Article ADS CAS PubMed Google Scholar * Strand, P. et al. Assessment of Fukushima-derived radiation doses and effects on wildlife in Japan. _Environ. Sci. Technol. Lett._ 1, 198–203
(2014). Article CAS Google Scholar * Kubota, Y. et al. Estimation of absorbed radiation dose rates in wild rodents inhabiting a site severely contaminated by the Fukushima Dai-Ichi
nuclear power plant accident. _J. Environ. Radioact_. 142, 124–131 (2015). Article CAS PubMed Google Scholar * The International Commission on Radiological Protection. Assessing effects
in terms of derive consideration levels in _environmental protection: the concept and use of reference animals and plants. ICRP publication 108. Annals of the ICRP_. (ed. Jalentin J) 88–96
(Pergamon, 2008). * Ohdachi, S. D., Ishibashi, Y., Iwasa, M. A. & Saitoh, T. _The wild Mammals of Japan_ (Shoukaido Book Sellers and the Mammalogical Society of Japan, 2009). * Oka, T.
Home range and mating system of two sympatric field mouse species, _Apodemus speciosus_ and _Apodemus argenteus_. _Ecol. Res._ 7, 163–169 (1992). Article Google Scholar * Roots, R. &
Okada, S. Protection of DNA molecules of cultured mammalian cells from radiation-induced single-strand scissions by various alcohols and SH compounds. _Int. J. Radiat. Biol. Relat. Stud.
Phys. Chem. Med._ 21, 329–342 (1972). Article CAS PubMed Google Scholar * Nikitaki, Z., Hellweg, C. E., Georgakilas, A. G. & Ravanat, J. L. Stress-induced DNA damage biomarkers:
applications and limitations. _Front. Chem._ 3, 35 (2015). Article ADS PubMed PubMed Central Google Scholar * Dizdaroglu, M. Mechanisms of oxidative DNA damage: lesions and their
measurement. In _Advances in DNA Damage and Repair: Oxygen Radical Effects, Cellular Protection, and Biological con-sequences_ (eds Dizdaroglu, M. & Karakaya, A. E.) 67–87 (Kluwer
Academic, 1999). * Slupphaug, G., Kavli, B. & Krokan, H. E. The interacting pathways for prevention and repair of oxidative DNA damage. _Mutat. Res._ 531, 231–251 (2003). Article CAS
PubMed Google Scholar * Loft, S. & Poulsen, H. E. Cancer risk and oxidative DNA damage in man. _J. Mol. Med._ 74, 297–312 (1996). Article CAS PubMed Google Scholar * Shibutani, S.,
Takeshita, M. & Grollman, A. P. Insertion of specific bases during DNA synthesis past the oxidation-damaged base 8-oxodG. _Nature_ 349, 431–434 (1991). Article ADS CAS PubMed Google
Scholar * Radicella, J. P., Dherin, C., Desmaze, C., Fox, M. S. & Boiteux, S. Cloning and characterization of hOGG1, a human homolog of the OGG1 gene of _Saccharomyces cerevisiae_.
_Proc. Natl. Acad. Sci. USA._ 94, 8010–8015 (1997). * Rosenquist, T. A., Zharkov, D. O. & Grollman, A. P. Cloning and characterization of a mammalian 8-oxoguanine DNA glycosylase. _Proc.
Natl. Acad. Sci. U S A_. 94, 7429–7434 (1997). Article ADS CAS PubMed PubMed Central Google Scholar * Karahalil, B., Hogue, B. A., De Souza-Pinto, N. C. & Bohr, V. A. Base
excision repair capacity in mitochondria and nuclei: tissue-specific variations. _FASEB J._ 16, 1895–1902 (2002). Article CAS PubMed Google Scholar * Risom, L., Moller, P., Vogel, U.,
Kristjansen, P. E. & Loft, S. X-ray-induced oxidative stress: DNA damage and gene expression of HO-1, ERCC1 and OGG1 in mouse lung. _Free Radic Res._ 37, 957–966 (2003). Article CAS
PubMed Google Scholar * Danielsen, P. H. et al. DNA damage in rats after a single oral exposure to diesel exhaust particles. _Mutat. Res._ 637, 49–55 (2008). Article CAS PubMed Google
Scholar * Li, D., Liu, Q., Gong, Y., Huang, Y. & Han, X. Cytotoxicity and oxidative stress study in cultured rat sertoli cells with methyl tert-butyl ether (MTBE) exposure. _Reprod.
Toxicol._ 27, 170–176 (2009). Article CAS PubMed Google Scholar * Okano, T. et al. Effects of environmental radiation on testes and spermatogenesis in wild large Japanese field mice
(_Apodemus speciosus_) from Fukushima. _Sci. Rep._ 6, 23601 (2016). Article ADS CAS PubMed PubMed Central Google Scholar * Pentreath, R. J. et al. ICRP publication 124: protection of
the environment under different exposure situations. _Ann. ICRP_. 43, 1–58 (2014). Article CAS PubMed Google Scholar * Zelko, I. N., Mariani, T. J. & Folz, R. J. Superoxide dismutase
multigene family: a comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) gene structures, evolution, and expression. _Free Radic Biol. Med._ 33, 337–349 (2002). Article CAS
PubMed Google Scholar * Rola, R. C., Marins, L. F., Nery, L. E., Rosa, D., Sandrini, J. Z. & C. E. & Responses to ROS inducer agents in zebrafish cell line: differences between
copper and UV-B radiation. _Fish. Physiol. Biochem._ 40, 1817–1825 (2014). Article CAS PubMed Google Scholar * Zhang, C. et al. Low-dose radiation induces renal SOD1 expression and
activity in type 1 diabetic mice. _Int. J. Radiat. Biol._ 90, 224–230 (2014). Article CAS PubMed Google Scholar * Minowa, O. et al. Mmh/Ogg1 gene inactivation results in accumulation of
8-hydroxyguanine in mice. _Proc. Natl. Acad. Sci. U S A_. 97, 4156–4161 (2000). Article ADS CAS PubMed PubMed Central Google Scholar * Klungland, A. et al. Accumulation of premutagenic
DNA lesions in mice defective in removal of oxidative base damage. _Proc. Natl. Acad. Sci. U S A_. 96, 13300–13305 (1999). Article ADS CAS PubMed PubMed Central Google Scholar * Ohno,
M. et al. 8-oxoguanine causes spontaneous de novo germline mutations in mice. _Sci. Rep._ 4, 4689 (2014). Article PubMed PubMed Central Google Scholar * Xie, Y. et al. Deficiencies in
mouse myh and Ogg1 result in tumor predisposition and G to T mutations in codon 12 of the K-ras oncogene in lung tumors. _Cancer Res._ 64, 3096–3102 (2004). Article CAS PubMed Google
Scholar * Maehata, I. Fauna of Muridae and the seasonal changes in the number and reproductive status of _Apodemus speciosus_ in the evergreen and deciduous broad-leaved forests (in
Japanese with English summary). _Bull. Cent. Nat. Environ. Educ. Nara Univ. Educ._ 1, 21–32 (1996). Google Scholar * Okano, T. et al. Classification of the spermatogenic cycle, seasonal
changes of seminiferous tubule morphology and estimation of the breeding season of the large Japanese field mouse (_Apodemus speciosus_) in Toyama and Aomori prefectures, Japan. _J. Vet.
Med. Sci._ 77, 799–807 (2015). Article PubMed PubMed Central Google Scholar * Takino, S. et al. Analysis of the effect of chronic and low-dose radiation exposure on spermatogenic cells
of male large Japanese field mice (_Apodemus speciosus_) after the Fukushima Daiichi nuclear power plant accident. _Radiat. Res._ 187, 161–168 (2019). Article ADS Google Scholar *
Dubrova, Y. E. et al. Transgenerational mutation by radiation. _Nature_ 405, 37 (2000). Article ADS CAS PubMed Google Scholar * Szumiel, I. Ionizing radiation-induced oxidative stress,
epigenetic changes and genomic instability: the pivotal role of mitochondria. _Int. J. Radiat. Biol._ 91, 1–12 (2015). Article CAS PubMed Google Scholar * Murakami, O. Growth and
development of the Japanese wood mouse (_Apodemus speciosus_). I. The breeding season in the field. _Jap J. Ecol._ 24, 194–206 (1974). Google Scholar * Committee of Reviewing Taxon Names.
Specimen collections in the Mammal Society of Japan. Guidelines for using wild mammals. _Mamm. Sci._ 49, 303–309 (2009). Google Scholar * Onuma, M., Endoh, D., Ishiniwa, H. & Tamaoki,
M. Estimation of dose rate for the large Japanese field mouse (_Apodemus speciosus_) distributed in the difficult-to-return zone in Fukushima prefecture in Low-dose Radiation Effects on
Animals and Ecosystems (ed Fukumoto, M.) 17–30 (Springer, (2020). * Plante, I. & Cucinotta, F. A. Cross sections for the interactions of 1 eV–100 MeV electrons in liquid water and
application to Monte-Carlo simulation of HZE radiation tracks. _New. J. Phys._ 11, 063047 (2009). Article ADS Google Scholar * Kishimoto, M. & Mori, Y. Body hair features of the
Japanese dormouse: comparison with the large Japanese field mouse and local variation. _Mamm. Sci._ 57, 61–68 (2016). Google Scholar * Todo, H. Transdermal permeation of drugs in various
animal species. _Drug Deliv Syst._ 32, 411–417 (2017). Article CAS Google Scholar * Hirayama, H., Namito, Y., Bielajew, A. F., Wilderman, S. J. & Nelson, W. R. _The EGS5 Code System.
SLAC Report Number: SLAC-R-730, KEK Report Number 2005-8_ http://rcwww.kek.jp/research/egs/egs5_manual/slac730-160113.pdf (2005). * Cook, C. S. & Langer, L. M. The beta-spectra of Cu64
as a test of the Fermi theory. _Phys. Rev._ 73, 601–607 (1948). Article ADS CAS Google Scholar * Hsue, S. T., Langer, L. M. & Tang, S. M. Precise determination of the shape of the
twice forbidden beta spectrum of 137Cs. _Nucl. Phys._ 86, 47–55 (1966). Article CAS Google Scholar * Oakberg, E. F. A description of spermiogenesis in the mouse and its use in analysis of
the cycle of the seminiferous epithelium and germ cell renewal. _Am. J. Anat._ 99, 391–413 (1956). Article CAS PubMed Google Scholar * Perey, B., Clermont, Y. & Leblond, C. The wave
of the seminiferous epithelium in the rat. _Am. J. Anat._ 108, 47–77 (1961). Article Google Scholar * Ishiniwa, H., Sogawa, K., Yasumoto, K. & Sekijima, T. Polymorphisms and
functional differences in aryl hydrocarbon receptors (AhR) in Japanese field mice, _Apodemus speciosus_. _Environ. Toxicol. Pharmacol._ 29, 280–289 (2010). Article CAS PubMed Google
Scholar * Schneider, G., Chicken, E. & Becvarik, R. _NSM3: Functions and Dataset to Accompany Hollander, Wolfe, and Chicken – Nonparametric Statistical Methods, Third Edition. R Package
Version 1.15_ http://CRAN.R-project.org/package=NSM3 (2020). * Kuznetsova, A., Brockhoff, P. B. & Christenson, R. H. B. lmerTest package: tests in linear mixed effects models. _J. Stat.
Softw._ 62, 1–26 (2017). Google Scholar * Bates, D., Mächler, M., Bolker, B. & Walker, S. Fitting linear mixed-effects models using lme4. _J. Stat. Softw._ 67, 1–48 (2015). Article
Google Scholar * Brooks, M. E. et al. glmmTMB balances speed and flexibility among packages for zero-inflated generalized liner mixed modeling. _R J._ 9, 378–400 (2017). Article Google
Scholar * Quinn, G. P., Keough, M. J. & Correlation and regression in _Experimental design and data analysis for biologist_ (eds. Quinn, G. P. & Keough, M. J.) 72–110 (Cambridge
University Press, 2002). * Huang, C. J. & Bolch, B. W. On the testing of regression disturbances for normality. _J. Am. Stat. Assoc._ 69, 330–335 (1974). Article Google Scholar * R
Core Team. _R: A Language and Environment for Statistical Cumputing. R Foundation for Statistical Computing._ (2019). https://www.R-project.org/ Download references ACKNOWLEDGEMENTS We thank
Dr. Yoshiki Tate for technical support with 8-OHdG immunostaining and Dr. Takehiko Hiwatari for assistance with the measurement of radioactive Cs in the samples. This study was financially
supported by the project “Study on the dynamics of radioactive materials in a multimedia environment” commissioned by the Ministry of the Environment, Japan. AUTHOR INFORMATION AUTHORS AND
AFFILIATIONS * Ecological Risk Assessment and Control Section, Center for Environmental Biology and Ecosystem Studies, National Institute for Environmental Studies, 16-2, Onogawa, Tsukuba,
Ibaraki, 305-8506, Japan Hiroko Ishiniwa & Manabu Onuma * Radioecologcial Transfer and Effects Division, Institute of Environmental Radioactivity, Fukushima University, 1 Kanayagawa,
Fukushima, Fukushima, 960-1296, Japan Hiroko Ishiniwa * Ecological Genetics Analysis Section, Center for Environmental Biology and Ecosystem Studies, National Institute for Environmental
Studies, 16-2, Onogawa, Tsukuba, Ibaraki, 305-8506, Japan Tsukasa Okano * Department of Radiation Biology, School of Veterinary Medicine, Rakuno Gakuen University, 582,
Bunkyodai-Midorimachi, Ebetsu, Hokkaido, 069-8501, Japan Daiji Endoh * High Energy Accelerator Research Organization, 1-1, Oho, Tsukuba, Ibaraki, 305-0801, Japan Hideo Hirayama *
Environmental Impact Assessment Section, Fukushima Regional Collaborative Research Center, National Institute for Environmental Studies, 10-2, Fukasaku, Miharu, Fukushima, 963-7700, Japan
Akira Yoshioka * Faculty of Science, Academic Assembly, University of Toyama, Gofuku 3190, Toyama, Toyama, 930-8555, Japan Yasushi Yokohata * Laboratory of Wildlife Science, School of
Veterinary Medicine, Kitasato University, 23-35-1, Higashi, Towada, Aomori, 034-8628, Japan Junji Shindo * Division of Bio-Resources, Department of Biotechnology, Frontier Science Research
Center, University of Miyazaki, 5200, Kihara, Kiyotake, Miyazaki, 889-1692, Japan Chihiro Koshimoto, Akio Shinohara & Shinsuke H. Sakamoto * Department of Animal and Grassland Sciences,
Faculty of Agriculture, University of Miyazaki, Gakuen-kibanadai-nishi-1-1, Miyazaki, 889-2192, Japan Shinsuke H. Sakamoto * Biodiversity Division, National Institute for Environmental
Studies, 16-2, Onogawa, Tsukuba, Ibaraki, 305-8506, Japan Masanori Tamaoki & Manabu Onuma Authors * Hiroko Ishiniwa View author publications You can also search for this author inPubMed
Google Scholar * Tsukasa Okano View author publications You can also search for this author inPubMed Google Scholar * Daiji Endoh View author publications You can also search for this author
inPubMed Google Scholar * Hideo Hirayama View author publications You can also search for this author inPubMed Google Scholar * Akira Yoshioka View author publications You can also search
for this author inPubMed Google Scholar * Yasushi Yokohata View author publications You can also search for this author inPubMed Google Scholar * Junji Shindo View author publications You
can also search for this author inPubMed Google Scholar * Chihiro Koshimoto View author publications You can also search for this author inPubMed Google Scholar * Akio Shinohara View author
publications You can also search for this author inPubMed Google Scholar * Shinsuke H. Sakamoto View author publications You can also search for this author inPubMed Google Scholar *
Masanori Tamaoki View author publications You can also search for this author inPubMed Google Scholar * Manabu Onuma View author publications You can also search for this author inPubMed
Google Scholar CONTRIBUTIONS H.I., T.O., M.O., Y.Y., and J.S. collected the specimens; H.I., T.O., M.O., K.C., A.S., and S.H.S. performed the experiments; D.E. and H.H. performed the EGS5
simulations; A.Y. performed the statistical analysis; M.O. and M.T. designed the study; and H.I. and M.O. wrote the paper. All authors contributed extensively to the discussion regarding
this work and reviewed the manuscript. CORRESPONDING AUTHOR Correspondence to Manabu Onuma. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. ADDITIONAL
INFORMATION PUBLISHER’S NOTE Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. ELECTRONIC SUPPLEMENTARY MATERIAL Below is
the link to the electronic supplementary material. SUPPLEMENTARY MATERIAL 1 SUPPLEMENTARY MATERIAL 2 SUPPLEMENTARY MATERIAL 3 RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed
under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or
format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material.
You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the
article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use
is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit
http://creativecommons.org/licenses/by-nc-nd/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Ishiniwa, H., Okano, T., Endoh, D. _et al._ Oxidative stress on the male
reproductive organs of wild mice collected from an area contaminated by radioactive materials in Fukushima. _Sci Rep_ 14, 29706 (2024). https://doi.org/10.1038/s41598-024-80869-2 Download
citation * Received: 09 December 2023 * Accepted: 22 November 2024 * Published: 29 November 2024 * DOI: https://doi.org/10.1038/s41598-024-80869-2 SHARE THIS ARTICLE Anyone you share the
following link with will be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer
Nature SharedIt content-sharing initiative



:max_bytes(150000):strip_icc():focal(666x0:668x2)/elvis-costello-2-54be89e8f2e04b25ac90b50c834899e0.jpg)