- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT Secondary metabolites (SMs) are the primary source of therapeutics and lead chemicals in medicine. They have been especially important in the creation of effective cures for
conditions such as cancer, malaria, bacterial and fungal infections, neurological and cardiovascular problems, and autoimmune illnesses. In the present study, _Aspergillus pseudodeflectus_
AUMC 15761 was demonstrated to use wheat bran in solid state fermentation (SSF) at optimum conditions (pH 7.0 at 30 °C after 10 days of incubation and using sodium nitrate as a nitrogen
source) to produce methyl ferulate and oleic acid with significant antioxidant and antibacterial properties. Gas chromatography-mass spectrometry (GC–MS) analysis of the crude methanol
extract revealed eleven peaks that indicated the most common chemical components. Purification of methyl ferulate and oleic acid was carried out by column chromatography, and both compounds
were identified by in-depth spectroscopic analysis, including 1D and 2D NMR and HR-ESI–MS. DPPH activity increased as the sample concentration increased. IC50 values of both compounds
obtained were 73.213 ± 11.20 and 104.178 ± 9.53 µM, respectively. Also, the MIC value for methyl ferulate against _Bacillus subtilis_ and _Staphylococcus aureus_ was 0.31 mg/mL, while the
corresponding MIC values for oleic acid were 1.25 mg/mL and 0.62 mg/mL for both bacterial strains, respectively. Molecular modeling calculations were carried out to reveal the binding mode
of methyl ferulate and oleic acid within the binding site of the crucial proteins of _Staphylococcus aureus_. The docking results were found to be well correlated with the experimental data.
SIMILAR CONTENT BEING VIEWED BY OTHERS A SCAFFOLDED APPROACH TO UNEARTH POTENTIAL ANTIBACTERIAL COMPONENTS FROM EPICARP OF MALAYSIAN _NEPHELIUM LAPPACEUM_ L. Article Open access 05 July
2021 MOLECULAR DOCKING ANALYSIS AND EVALUATION OF THE ANTIMICROBIAL PROPERTIES OF THE CONSTITUENTS OF _GERANIUM WALLICHIANUM_ D. DON EX SWEET FROM KASHMIR HIMALAYA Article Open access 22
July 2022 METABOLITE FINGERPRINTING OF PHYTOCONSTITUENTS FROM _FRITILLARIA CIRRHOSA_ D. DON AND MOLECULAR DOCKING ANALYSIS OF BIOACTIVE PEONIDIN WITH MICROBIAL DRUG TARGET PROTEINS Article
Open access 04 May 2022 INTRODUCTION Currently, infectious diseases related to bacteria, fungi, and viruses represent a significant issue for global health1. Worldwide, acute respiratory
infections (ARIs), intestinal infections, HIV/AIDS, tuberculosis, and malaria annually account for roughly 4, 3, 1.8, 0.7, and 1.3 million deaths, respectively2,3. Rising microbial
resistance to conventional medication has had an significant impact on research into novel substances that demonstrate broad-spectrum antibacterial activity4. Secondary metabolites are
synthesized by a range of plants, animals, and microorganisms. Fungi, which are particularly prolific sources of bioactive secondary metabolites5, continue to be among the most important
organisms investigated for therapeutic agents and lead compounds in medicine. They have been especially important in the development of effective therapies for cancer, malaria, bacterial and
fungal infections, neurological and cardiovascular diseases, and autoimmune disorders6. Fungi create a broad array of secondary metabolites, some of which are harmful to humans, plants,
animals, and the environment7. In contrast, they also serve as a reliable source for the creation of a number of beneficial products, including enzymes8,9,10, biodiesel11, fatty acids12, and
mycotoxins13, as well as being used to make pharmaceutical, agrochemical, and cosmetic commodities. Despite the fact that many secondary metabolites generated from fungi have been
discovered previously, there are still many others that have yet to be discovered. Only a few secondary metabolites have been recognized from fungi, although there have been more than
species of 150,000 fungi identified thus far (Species Fungorum database). This is because it is difficult to find and recognize new secondary metabolites7. Methyl ferulate, which was first
detected from the medicinal plant _Stemona tuberosa_, has the ability to cross cell membranes and enter the brain; it also exhibits anti-free radical characteristics14,15. Methyl ferulate
has also been found in a diverse range of fruits such as oranges and tomatoes, and in some cereals such as rice and corn16,17,18,19. Because of its low toxicity and diminishing oxidation
activity, it has been utilized effectively as a food additive20,21 in cosmetics, as well as in health care and skin care products14. Oleic acid is a natural product that has recently become
commonly used to prevent food from oxidizing and it also has great antimicrobial potential value against many fungi and bacteria22,23. Oleic acid, which makes up around 80% of the total
fatty acids in virgin olive oil24, is used to decrease cholesterol and reduce inflammation in order to avoid heart diseases25,26,27. Globally, there is a huge accumulation of agricultural
and industrial waste. Nevertheless, because these wastes usually include a substantial content of carbohydrates, minerals, and proteins along with cellulose (30–40%), hemicellulose (20–40%),
and lignin (20–30%)28, they should not be regarded as "wastes" but rather as "raw materials" for other industrial processes29. Notwithstanding the fact that they are
currently underutilized in Egypt, a variety of agro-industrial wastes have been employed as substrates in the solid state fermentation process due to their availability, low cost,
environmental friendliness, prolonged shelf life, and simplicity of downstream processing30,31. As a result, the present study focused on the production of methyl ferulate and oleic acid by
_Aspergillus pseudodeflectus_ AUMC 15761 from wheat bran under solid state fermentation conditions as well as on methods to purify, identify, and employ these metabolites as antioxidant and
antibacterial agents. In addition, the binding mode of the two obtained compounds within the binding site of the crucial proteins of _Staphylococcus aureus_ was investigated using molecular
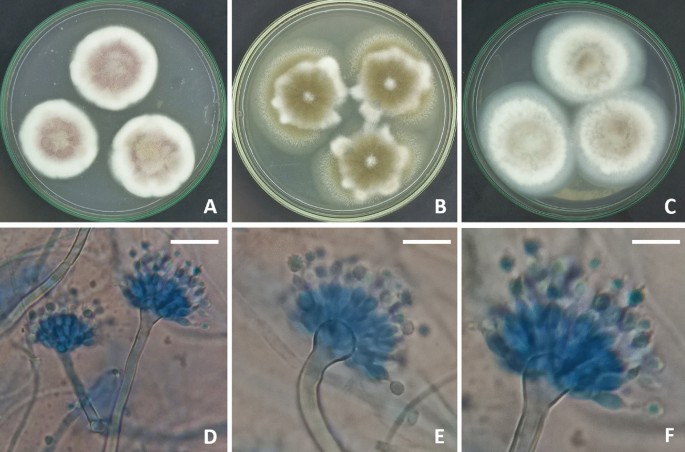
modeling computations. RESULTS MORPHOLOGICAL AND MOLECULAR IDENTIFICATION OF _ASPERGILLUS PSEUDODEFLECTUS_ STRAIN The morphological characteristics of the _Aspergillus_ strain used in this
study shared the identical features of _A. pseudodeflectus_ as having radiate, brown conidial heads. Stipes 35–200 × 2.5–3.5 µm. Vesicles globose to clavate, 4–12 µm. Conidia globose to
ellipsoidal, brown, 3.5–5 µm. Hülle cells absent (Fig. 1). Phylogenetic analysis based on ITS sequencing was employed to confirm the identification of the strain. The final ITS data set
contained 20 sequences that produced 616 characters, of which 505 characters could be correctly aligned, 46 characters were counted as variable, and 8 as informative. The Tamura 3-parameter
using a discrete Gamma distribution (T92 + G) was the perfect model used to represent the relationship among taxa. The maximum Parsimony method yielded 10 trees, the most parsimonious of
which (Fig. 2) has a tree length of 61, the highest log likelihood of − 1180.74, consistency index of 0.733333, retention index of 0.818182, and a composite index of 0.600000 is shown in
Fig. 2. PRODUCTION OF SECONDARY METABOLITES BY _ASPERGILLUS PSEUDODEFLECTUS_ AUMC 15761 UTILIZING LIGNOCELLULOSIC WASTES The five lignocellulosic wastes (barley bran, date palm leaves,
orange peels, rice straw, and wheat bran) used in SSF were fermented in different proportions by _A. pseudodeflectus_ AUMC 15761. Wheat bran produced the most powerful crude extract that
exhibited the largest inhibition zone against the examined strains. Following _Escherichia. coli_ in terms of severity of impact were _Bacillus subtilis_, _Staphylococcus aureus_, and
_Staphylococcus epidermidis_ (Fig. 3). GC–MS ANALYSIS The GC–MS analysis of the methanol extract was carried out to evaluate its potential components since wheat bran extract was determined
to be the most promising. Based on the retention time, molecular weight, and fragmentation pattern of the most prominent chemicals, the current results revealed eleven peaks. These were
shown to have retention times of 23.222, 27.429, and 29.769 min, respectively, for _trans_-Ferulic acid, 3-(2, 5-Dimethoxyphenyl) propionic acid, and oleic acid (Table 1; Fig. 4).
OPTIMIZATION OF PRODUCTION CONDITIONS OF THE BIO-ACTIVE SECONDARY METABOLITES USING WHEAT BRAN Based on a one factor at a time (OFAT) analysis, the results obtained revealed that _A.
pseudodeflectus_ AUMC 15761 could produce the most bio-active secondary metabolites with the greatest effect against the tested bacteria at pH 7.0 using sodium nitrate as a nitrogen supply
after 10 days of incubation at 30 °C. These optimal conditions were found to cause the greatest inhibition of the four tested bacterial strains. The inhibition zones were 20.5 ± 1.3, 37.9 ±
1.7, 27.0 ± 1.8, and 19.0 ± 1.7 mm for _B. subtilis_, _E. coli_, _S. aureus_, and _S. epidermidis_, respectively (Fig. 5). PRODUCTION AND PURIFICATION OF BIO-ACTIVE SECONDARY METABOLITES BY
COLUMN CHROMATOGRAPHY _Aspergillus pseudodeflectus_ AUMC 15761 could ferment 500 g of wheat bran in SSF, and 90.0 g (18%) of crude extract were produced. Using _n_-hexane, dichloromethane
(DCM), and 0–100% gradients of MeOH in the DCM solvent system, twelve fractions (F1–F12) were obtained after fractionation by means of the VLC column (Table 2), Fraction F3 (750 mg), which
was eluted by DCM: MeOH (95: 5) had the highest inhibition (23.0 ± 1.8, 32.9 ± 1.6, 42.5 ± 1.4, and 39.4 ± 1.5 mm) against _B. subtilis_, _E. coli_, _S. aureus_, and _S. epidermidis_,
respectively. Consequently, it was subjected to purification by silica gel open column (1 × 100 cm). Nine sub-fractions (F3S1–F3S9) were subsequently produced as a result, with F3S8 (40 mg)
and F3S4 (100 mg) being the two most active. Further purification using a 0.5 × 25 cm open column of both fractions, produced a pure compound 2 (40 mg) from F3S4, while F3S8 required a final
purification step by preparative TLC plates (60 PF254) to obtain the pure compounds 1 (8.0 mg). HR-ESI MS AND NMR SPECTROSCOPIC ANALYSIS Compound 1 was isolated from _A. pseudodeflectus_
AUMC 15761 as an off-white powder (8.0 mg), suggesting a molecular formula of C11H12O4 as deduced from its HR-ESI–MS spectrum (Fig. 6) which exhibited a [M−H]− peak at _m/z_ 207.0664 and a
[M−H + H2O]− peak at _m/z_ 224.9995. The APT NMR spectrum of 1 (Table 3; Fig. 7), along with the HSQC analysis, confirmed the presence of 11 carbon atoms. These showed four quaternary
carbons (including one carbonyl ester resonates at δC 167.8), five methines (including an olefinic double bond at δC 145.1 and 115.3), and two methoxy methyls (δC 51.7 and 56.0). The 1H NMR
spectrum of 1 (Table 3; Fig. 8), confirmed the presence of an olefinic double bond at δH 7.63 (1H, d, J = 16.0 Hz) and δH 7.28 (1H, d, J = 16.0 Hz). The HSQC correlation between the rest
three methines and their corresponding protons [δC 109.9/δH 7.02 (1H, d, J = 2.0 Hz), δC 114.8/δH 6.91 (1H, d, J = 8.0 Hz), and δC 123.1/δH 7.06 (1H, dd, J = 2.0, 8.0 Hz)], along with the
rest three quaternaries, revealed the presence of a tri-substituted benzene ring (Table 3; Fig. 9). The olefinic proton resonates at δH 7.28 and showed 1H-1H cosy correlations to the
olefinic proton at δH 7.63 (Table 3; Fig. 10) and HMBC correlations with both the carbonyl carbon at δC 167.8 and the aromatic carbon at δC 127.1, indicating that 1 is a cinnamic acid
derivative (Table 3; Fig. 11). The HMBC correlations between the methoxy protons [δH 3.79 (3H, s)] and the carbonyl carbon (δC 167.8) confirmed that 1 is a cinnamic acid methyl ester
derivative. The other methoxy group at δH 3.92 (3H, s) showed a HMBC correlation with the quaternary aromatic carbon δC 146.9, and was downfield of the quaternary carbon δC 148.1, indicating
that 1 is a ferulic acid methyl ester. The stereochemistry of the double bond was determined to be trans by the large coupling constant of its olefinic protons [δH 7.28 (1H, d, J = 16.0
Hz), at δH 7.63 (1H, d, J = 16.0 Hz)]. Based on this cumulative analysis, the structure of 1 was established as methyl ferulate (Fig. 12). Compound 2 was isolated from _A. pseudodeflectus_
AUMC 15761 as a colorless oil (40.0 mg). HR-ESI–MS (Fig. 13) suggested the molecular formula as C18H34O2 according to the mass peak [M + H]+ at _m/z_ 283.2641. The 1H-NMR spectrum (Table 4;
Fig. 14) exhibited a protons signal at δH 5.34 (2H, m, H-9, H–10) and δH 2.01 (4H, m, H–8, H–11), assignable to olefinic protons and allylic protons, respectively. The spectrum also revealed
a methylene group α to carbonyl functionality (δH 2.34, 2H, t, J = 7.6, H-2). This was also substantiated by the APT NMR spectrum of 2 (Table 4; Fig. 15) which revealed the presence of a
carbonyl carbon signal (δC 180.4, C–1), two olefinic carbon signals (δC 130.2 and 129.9, C–9, C–10), a group of methylene carbons resonances at δC 22.8–34.2, and finally a primary methyl
group signal (δC 14.3, C–18), all of which were in agreement with a monounsaturated fatty acid. Thus, by comparing the 1H, 13C-NMR, and mass data of compound 2, oleic acid was therefore
determined to be the substance involved (Fig. 12). ANTIOXIDANT ACTIVITY OF METHYL FERULATE AND OLEIC ACID PRODUCED BY _ASPERGILLUS PSEUDODEFLECTUS_ AUMC 15761 The results of the current
study revealed two antioxidant substances—methyl ferulate and oleic acid. The DPPH activity increased as the sample concentration increased (Fig. 16A), with IC50 values of 73.213 ± 11.20 and
104.178 ± 9.53 µM were significantly greater (_p_ < 0.05) respectively, for both substances, as compared to the value (60.299 ± 4.769 µM) of ascorbic acid (Fig. 16B; Table 5).
ANTIBACTERIAL ACTIVITY OF METHYL FERULATE AND OLEIC ACID In the present study, the antibacterial tests against _B. subtilis_ and _S. aureus_ demonstrated a significant antibacterial impact
for both methyl ferulate and oleic acid. The MIC of methyl ferulate against the two bacterial strains was 0.31 mg/mL, while the MICs of pure oleic acid against both strains were 1.25 and
0.62 mg/mL, respectively (Tables 6, 7; Fig. 17). _R_ = zero (no linear association between the variables or no consistent linear component to that relationship); _R_ = 1 (perfect positive
linear relationship between the variables); 0 < _R_ < 1 (positive linear trend and the sampled individuals are scattered around the line of best fit; the smaller the absolute value of
_R_ the less well the data can be visualized by a single linear relationship. M = mean; %MB = mean value divided by the standard value. DOCKING COMPUTATIONS In order to recognize target
proteins for the antibacterial activity of the two compounds obtained in the present study, four various protein targets of _Staphylococcus aureus_ were investigated, including dihydrofolate
reductase (PDB code: 5ISP32), pyruvate kinase (PDB code: 5OE333), and sortase A (PDB code: 2MLM34). Before generation of data, the performance of the AutoDock 4.2.6 software was evaluated
via the re-docking of the co-crystalized inhibitors towards their targets. Based on the data illustrated in Fig. 18, the anticipated binding modes were essentially identical to their native
structures with RMSD values less than 1.0 Å, displaying the strong accuracy of the utilized technique. Using the assessed protocol, the docking pose of oleic acid and methyl ferulate with
dihydrofolate reductase, pyruvate kinase, and sortase A was predicted and presented in Fig. 18. As illustrated in Fig. 19, oleic acid and methyl ferulate exhibited multiple H-bonds and
additional interactions involving vdW (van der Waals), pi-based, and hydrophobic interactions. More precisely, oleic acid displayed an eminent docking score towards dihydrofolate reductase
compared to methyl ferulate, with values of − 6.3 and − 5.7 kcal/mol, respectively (Table 8). Inspection the binding mode of oleic acid demonstrated that 2 H-bonds with GLN19 (1.78 Å) and
LEU20 (2.18 Å). However, methyl ferulate exhibited 3 H-bonds with SER49 (1.68 Å), ALA7 (1.98 Å), and TYR98 (2.44 Å). For pyruvate kinase, oleic acid, and methyl ferulate, the exposed docking
scores had values of − 6.3 and − 5.7 kcal/mol, respectively. Oleic acid formed a hydrogen bond with GLY307 (2.10 Å). In comparison, methyl ferulate exhibited four hydrogen bonds with HIS394
(2.26 Å), GLY302 (2.01 Å), GLY279 (1.88 Å), and GLY300 (2.16 Å). Sortase A, oleic acid and methyl ferulate displayed good docking scores with values of − 5.2 and − 4.9 kcal/mol,
respectively. Oleic acid exhibited three hydrogen bonds with THR106 (1.99 Å), ARG139 (2.97 Å), and TRP136 (1.90 Å). On the other hand, methyl ferulate established two H-bonds with GLY61
(2.12 Å) and CYS126 (3.03 Å). DISCUSSION Secondary metabolites created by fungi have been shown to be a wonderful source of new drugs, biofuels, industrial chemicals, food additives, and
feed additives35. Penicillins, lovastatin, echinocandin B, and cyclosporine A offer examples of how important it is to investigate fungal sources for novel pharmaceuticals36. Aromatic
compounds, amino acids, fatty acids, butanolides, butenolides, cytochalasans, macrolides, naphthalenones, pyrones, and terpenes are only a few of the structural types of metabolites that
fungi produce36,37. There have been 15600 fungal metabolites identified from species of _Alternaria_, _Aspergillus, Claviceps_, _Fusarium_, and _Penicillium_38. In the present study, _A.
pseudodeflectus_ AUMC 15761 was identified based on sequencing of the ITS region. The fungus utilized wheat bran as a substrate in SSF to produce, for the first time, methyl ferulate and
oleic acid, as confirmed by an antibacterial bioassay. Methyl ferulate was first reported from the plant _Stemona tuberosa_14,15 and then re-isolated from _Coriolopsis aspera_39,40, _Morinda
citrifolia_41, and _Kigelia. africana_42. The _Aspergillus ustus_ group has been found to produce several active metabolites such as the three novel isoquinoline alkaloids, TMC-120A-C (1–3)
(furo [3, 2-h] isoquinoline-type) discovered from _A. ustus_ TC 111843; averufin, versicolorine C, along with austalides O and J have been isolated from different strains of _A. ustus_44; A
newly isochroman derivative, pseudodeflectusin (9-hydroxy-7-methyl-2-(methylethylidine)-furano [3, 2-H] isochroman-3-one), was discovered from _Aspergillus pseudodeflectus_45; Drimane
sesquiterpenoids 1, 15 were isolated from _Aspergillus pseudodeflectus_ strains F-1860, F-186144; Ophiobolin K and 6-epi-ophiobolin K, have been developed from _A. calidoustus_ UFMGCB
410746; Drimane sesquiterpenoids 1, 2, 7, and 15, TMC-120A-C, desferritriacetylfusigen and TMC-120 derivative 1, have been found in several strains of _A. calidoustus_44. In addition,
several bio-active secondary metabolites have been produced by many species of fungi in SmF or SSF47, such as penicillins and cephalosporins isolated from _Penicillium_ and _Acremonium_,
respectively48,49; feruloyl esterase produced by _Aspergillus terreus_ GA2 using maize bran50; methyl ferulate from the fruiting bodies of _Coriolopsis aspera_40; festuclavine 2 produced by
_A. fumigatus_51; cyclosporine 41, which exhibits broad spectrum of antifungal activity52, derived from _Toplypocladium inflatum_53; geranylgeraniol, farnesol, hexacosanol, oleic acid, and
squalene synthesized by _Colletotrichum coccodes_54; 1-Octacosanol produced by _Phyllosticta capitalensis_55; a set of peptaibols, that are extremely potent growth inhibitors of several
species of fungi, including the plant pathogens _Alternaria alternata_, _Phoma cucurbitaceum_, _Fusarium_ spp., as well as the human pathogen _A. fumigatus_, have been reported from
_Trichoderma reesei_56; and Echinocandin B 38 from _Aspergillus nidulans_57. The most potent antibacterial crude metaboliate from _Aspergillus pseudodeflectus_ AUMC 15761’s was produced
after 10 days of fermentation utilizing wheat bran in SSF at pH 7.0 and 30 °C, using sodium nitrate as a nitrogen source. The pH of the growth medium and other physical factors, such as the
incubation temperature, were found to have a substantial impact on the production of secondary metabolites, with synthesis rapidly decreasing on either side of an optimal point. By changing
the degree of dissociation of different molecules in the media, the quantity of hydrogen or hydroxyl ions may have direct or indirect effects on a cell. As a result, variations in pH have an
effect on the solubility and dissociation of intermediate products as well as the activity of microbial enzymes58,59. One of the unsaturated fatty acids formed by the reaction of palmitic
and stearic acids is oleic acid. Moreover, enzymatic activity can convert saturated fatty acids such as stearic acid and palmitic acid into oleic acid60. In the present study, the methanolic
extract of wheat bran fermented by _A. pseudodeflectus_ AUMC 15761 was utilized to isolate oleic acid. Saturated fatty acids and monounsaturated fatty acids, including palmitic and oleic
acids, occurred in abundance in the fatty acid profiles of _Mucor circinelloides_ URM 4140, _M. hiemalis_ URM 4144, and _Penicillium citrinum_ URM 412661. Oleic acid, also known as
9-octadecenoic acid, is a healthy kind of omega-9 unsaturated fatty acid that is very useful for people's health22. Unsaturated fatty acids do indeed lower cholesterol by activating
cholesterol acetyltransferase, as is widely known. Cancer, cardiovascular, autoimmune, Parkinson's, Alzheimer's, inflammatory, and hypertensive illnesses are all treated
effectively with fatty acids. These compounds have been employed as an anticancer treatment because they may cause cancer cells to undergo apoptosis and regulate the cell membrane22.
_Aspergillus terreus_, _Claviceps purpurea_, _Tolyposporium_ sp., _Mortierella alpina_, and _Mortierella isabellina_ are a few examples of species of fungi that may collect lipids. Although
several fungi may produce lipids, the majority of fungi are studied primarily for their capacity to create specific lipids such as docosahexaeneoic acid (DHA), gamma-linolenic acid (GLA),
eicosapentaenoic acid (EPA), and arachidonic acid (ARA)62. Bio-guided isolation of the secondary metabolites of the methanol extract of _A. pseudodeflectus_ AUMC 15761 led to the
purification of two active compounds—methyl ferulate and oleic acid. Their structures were determined by comparing their NMR and HR-EI-Ms data with that available from the
literature15,39,42. It is noteworthy that this is the first report of producing oleic acid and methyl ferulate from _A. pseudodeflectus._ Methyl ferulate and oleic acid in this study were
found to have a significant antibacterial potential against gram positive bacteria, showing the best activity against _B. subtilis_ and _S. aureus_, while _E. coli_ and _S. epidermidis_ were
not affected by either compound in the present study. In the study, the MICs for methyl ferulate and oleic acid against _B. subtilis_ and _S. aureus_ were 79% and 69%, respectively,
compared to the bacitracin standard, with MICs of 79% and 69%, and 58% and 40%, respectively. Similarly, the antibacterial activity of methyl ferulate against _Shigella putrefaciens_ was
determined63. Certain oils, including rose essential oil, have been proven to have antibacterial properties against _S. aureus_, and the efficacy against gram positive _S. aureus_ was
observed to be less than that of rifampin and gentamicin, with negligible MIC values64. Because of the observed sensitivity of gram-positive bacteria to the presence of one phenolic hydroxyl
group in methyl ferulate, its antibacterial mechanism was taken into consideration42,65. Although the exact mechanism of the antibacterial activity of fatty acids is unknown, it is thought
that their functional nature is connected to the permeability, membrane disruption, and fatal changes in the bacterial cytoplasm. As a result, rupture or alteration of the membrane-dependent
conduction systems may occur22,66._ Escherichia coli_, a normally resistant bacterium, becomes very susceptible to the bactericidal effects of fatty acids if the lipopolysaccharide outer
membrane is destroyed using ethylenediaminetetraacetic acid. As gram-negative bacteria, they are protected by their outer lipid membrane from the corrosive effects of oleic acid67. Methyl
ferulate has been reported to have antioxidant activity (% DPPH), with IC50 values of 73.213 11.20 µM, respectively41. Considering that, the principal mode of action of phenolic antioxidants
is believed to be the scavenging of free radicals68. Due to the presence of one phenolic hydroxyl group in methyl ferulate, its antioxidant mechanism was taken into consideration42,65.
Antioxidant activity (% DPPH) of OA with IC50 values of 104.178 ± 9.53 µM has been reported in the literature69. Methy ferulate is a new natural antibacterial agent with strong efficacy and
low toxicity. It has great potential for applications in food preservation63. Oleic acid, which accounts for about 80% of the total fatty acids in virgin olive oil, has recently become an
often used substance to protect food from oxidizing24. Oleic acid and methyl ferulate both had positive docking results against several _Staphylococcus aureus_ protein targets, based on the
docking results. The development of hydrogen bonding interactions with the active site of the _Staphylococcus aureus_ target proteins under investigation may be responsible for the high
docking scores of these two compounds. These findings shed additional light on the significance of the chemicals that have been identified as potential candidates for antibacterial
medication. MATERIALS AND METHODS MATERIALS AND CHEMICALS For the extraction, fractionation, and column chromatography, the organic solvents used were supplied by El-Nasr Pharmaceutical and
Chemical Co. (ADWIC), Egypt. The deuterated chloroform (CDCl3) used for NMR analysis was purchased from Sigma-Aldrich. TLC pre-coated plates (F254 & PF254) and silica gel for column
chromatography (70–230 and 230–400 Mesh) were provided by Merck (Darmstadt, Germany). STRAIN ISOLATION AND PRESERVATION Using the dilution plate technique70, the strain for this
investigation was isolated from a soil sample collected from Egypt's Aswan Governorate. Before adding Czapek's Dox agar (CzA) to Petri plates, the soil solution was appropriately
diluted. The cultures were then maintained for two weeks at 25 °C. To create pure cultures of the fungus, the developed colonies were purified on CzA utilizing the single spore isolation
technique71. MORPHOLOGICAL AND MOLECULAR IDENTIFICATION OF THE _ASPERGILLUS_ STRAIN For morphological identification of the strain of _Aspergillus_, the fungus was inoculated on Malt Extract
Agar (MEA), Czapek’s Yeast Autolysate Agar (CYA), and CzA72, and incubated for 7 days at 25 °C. The fungus in this study was morphologically identified on the basis of its macroscopic and
microscopic characteristics, following the relevant key of _Aspergillus_: section Usti73. This strain was deposited and designated as AUMC 15761 in the culture collection of the Assiut
University Mycological Centre. DNA isolation was carried out74, and the PCR reaction was performed by SolGent Co., Ltd (Daejeon, South Korea) using SolGent EF-Taq and the universal primers
ITS1 and ITS475. DNASTAR (version 5.05) was used to generate the contiguous sequences of the species of _Aspergillus_ used in this investigation. There are 20 sequences in the overall ITS
dataset that were used for phylogenetic analysis, consisting of one outgroup sequence for _Aspergillus creber_ NRRL 58,592 (NR_135442), the sequence for _Aspergillus pseudodeflectus_ (AUMC
15761 in this manuscript), and 18 sequences from the genus _Aspergillus_: section Usti acquired from GenBank. MAFFT (version 6.861b)76 with the default settings was used to align all
sequences. Optimization of the alignment gaps and sparse uninformative characters was conducted by BMGE77_._ Maximum-likelihood (ML) and maximum-parsimony (MP) phylogenetic analyses were
carried out using MEGA X (version 10.2.6)78, and 1000 replications79 were employed to assess the robustness of the most parsimonious trees. The ideal nucleotide substitution model for ML
analysis was identified using Modeltest 3.7's Akaike Information Criterion (AIC)80. After editing, the tree was saved in TIF format81_._ INOCULUM _Aspergillus pseudodeflectus_ AUMC
15761 was grown for 7 days on MEA at 25 °C and a spore solution (prepared in 10% tween 80) containing 1.5 × 108 spores/mL was used to inoculate the substrate (5 mL/10 g substrate)82. SOLID
STATE FERMENTATION (SSF) OF LIGNOCELLULOSIC WASTES In order to determine how the selected isolate of _A. pseudodeflectus_ produced an antibacterial chemical, five samples of agricultural
waste were studied. The fermentation materials included barley bran (BB), date palm leaves (DPL), orange peels (OP), rice straw (RS), and wheat bran (WB). They were purchased from local
marketplaces in Egypt's Assiut Governorate. Before being reduced to a size of 500 µm, they were cleaned with tap water to get rid of dirt and other impurities. As part of the
pretreatment procedure, the samples were treated with 1.0% NaOH, thoroughly filtered, and then washed with tap water. They were then dried at 60 °C83. In order to determine how the selected
_A. pseudodeflectus_ produced an antibacterial chemical, five samples of agricultural waste were studied. Separate Erlenmeyer flasks (500 mL) were filled with 10 g of the lignocellulosic
material, and the residue was then wetted down by 88% with a citrate buffer (pH 5.0). The flasks were next incubated for seven days at 30 °C. EXTRACTION OF BIO-ACTIVE COMPOUNDS Following the
incubation time, the flask contents underwent 48 h of oven drying at 60 °C. The mycelial mat and solid substrate were stirred in 50 mL of methanol for 2 h at 180 rpm in each flask. The
clear supernatant was obtained after centrifugation (10,000 rpm at 4 °C for 10 min). The volume of the methanol extract was then reduced by a rotary evaporator (Heidolph: Model reddot winner
2020; Germany). The sample was lyophilized into a powder using a freeze dryer (VirTis: Model 6 KBTES-55, NY; USA)84. ANTIBACTERIAL EFFECT OF THE CRUDE EXTRACTS The agricultural
waste-derived crude extract residue from each sample was dissolved in dimethyl sulfoxide (DMSO) at a 10% concentration. The antibacterial efficacy of the crude extract was assessed using the
agar well diffusion technique85, using 50 µL in each 5 mm well for _Escherichia coli_ ATCC 8739, _Bacillus subtilis_ ATCC 6633, _Staphylococcus aureus_ ATCC 6538, and _Staphylococcus
epidermidis_ ATCC 12228. Bacitracin (10 U) and Piperacillin/Tazobactam 10: 1 (110 µg/disc), served as a positive control. This test was performed after each stage of purification. GC–MS
ANALYSIS This analysis was carried out at the Analytical Chemistry Unit (ACAL), Faculty of Science, Assiut University, Egypt. Before being injected into a GC–MS device (7890A-5975B; Thermo
Scientific GC/MS; model ISQ; USA), with a nonpolar HP-5MS Capillary Standard column (30 × 0.25 × 0.25) mm, 0.5 g of the sample residue was dissolved in 5 mL of methanol and centrifuged for
15 min (10,000 rpm and 5 °C). The following was the cycle's parameters: oven program on at 120 °C for 5 min, 30 °C/min rising to 265 °C for 25 min, then 50 °C/min increased to 280 °C
for 5 min; run duration 48 min; post run 260 °C for 2 min; flow program 0.5 mL/min. for 10.9 min., and then 1 mL/min for 30 min. Equilibration time was 0.5 min, and the maximum temperature
280°C84. OPTIMIZATION OF BIO-ACTIVE SECONDARY METABOLITES PRODUCTION BY _A. PSEUDODEFLECTUS_ AUMC 15761 For maximization of secondary metabolites production, the respective pH (4.0, 5.0,
6.0, 7.0, and 8.0), temperature (20, 25, 30, 35, and 40 °C), nitrogen source (peptone, yeast extract, sodium nitrate, ammonium chloride, and ammonium sulphate, each at 0.2%), incubation time
(2, 4, 6, 8, up to 14 days) were adjusted using one factor at a time (OFAT)84. For the testing, a 10 g amount of wheat bran was placed in 500 mL Erlenmeyer flasks. Following the incubation
time, the bio-active Secondary metabolites were extracted as described above and then used in further tests. PRODUCTION OF BIO-ACTIVE SECONDARY METABOLITES BY _A. PSEUDODEFLECTUS_ AUMC 15761
UTILIZING WHEAT BRAN IN SSF Ten g portions of wheat bran were separately placed into 500 mL Erlenmeyer conical flasks, and the spore suspension (1.5 × 108 spore/mL) was prepared from a
seven-day-old _A. pseudodeflectus_ AUMC 15761 culture and added to each flask in a volume of 5.0 mL were used to inoculate the residue. The flasks were then kept under optimal production
cultural conditions. Following the incubation time, the flask contents were extracted, and the dried extract was applied throughout the purification process. PURIFICATION OF THE SECONDARY
METABOLITES BY COLUMN CHROMATOGRAPHY SAMPLE PREPARATION FOR COLUMN CHROMATOGRAPHY The residue was combined with an equivalent weight of silica gel powder and a trace amount of methanol was
added to create a slurry prior to each step of purification. The extract was placed onto a vacuum liquid chromatography (VLC) column for fractionation after being dried and slurred. The last
stage of purification was carried out by adding the obtained fractions to silica gel column chromatography84. VACUUM LIQUID CHROMATOGRAPHY (VLC) With 900 g of silica gel (230–400 Mesh), the
entire crude extract was fractionated using a VLC column (5.0 × 120 cm). The _n_-hexane, Dichloromethane (DCM) and 0–100% gradients of DCM in MeOH (by adding 10.0% MeOH each time) were used
in the fractionation process. A low-pressure evaporator was used to dry out the solvents after collecting a total of 250 mL of elutes. Fractions that produced the strongest antibacterial
properties and have comparable spots were combined, condensed, and dried for use in the further purification process. THIN LAYER CHROMATOGRAPH (TLC) TLC was carried out on pre-coated silica
gel F254 plates. A series of solvents of increasing polarities were used for developing the spots. For visualization of the spots, the plates were subjected to a UV inspection (at 365 and
254 nm) and then sprayed with 10% (v/v) H2SO4 in methanol, dried using a hot air drier, and heated to 110°C84. FINAL PURIFICATION OF THE ACTIVE SECONDARY METABOLITES The fraction containing
active chemicals was subsequently chromatographed on an open column (1.0 × 100 cm) equipped with a 35 g silica gel (70–230 Mesh). It was eluted in _n_–hexane gradients with 0–20% EtOAc
(adding 1.0% EtOAc each time). TLC was used to detect 25 mL elutes using three mobile systems (_n_–hexane: EtOAc: 95: 5, 90: 10, and 80: 20). After combining and drying similar elutes, nine
subfractions were obtained. The subfractions containing the most active compounds were uploaded over a second column (0.5 × 25 cm) packed with 15 g silica gel and a solvent system of
n–hexane: acetone (95: 5) was used for elution. A 15 mL elutes were collected, subjected to TLC, and those containing nearly pure compounds were combined. Following the method of Siddiqui,
et al.86, they were finally purified using preparative TLC plates (60 PF254). The antibacterial test for the purified compound was performed as described above. SPECTROSCOPIC NMR The
analysis was completed at the Micro-Analytical Unit (MAU) of Cairo University's School of Pharmacy in Giza, Egypt. Bruker Avance III HD 400 and 100 MHz spectrometers (Bruker Biospin,
Rheinstetten, Germany) and NMR software Topspin 3.2 pl 6 were used to produce the 1H and 13C-NMR spectra. The internal reference standard was tetramethylsilane (TMS). An LTQ Orbitrap XL
spectrometer was used to obtain the HR-ESI–MS data (Thermo Fisher Scientific; USA). DPPH RADICAL SCAVENGING ACTIVITY Using the methods described by Yen and Duh87, a freshly prepared methanol
solution containing 0.004% (w/v) of the 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical was refrigerated at 10 °C in the dark. Pure compound concentrations of 100, 50, 25, 12.5, 6.25, 3.125,
1.56, and 0.78 µM methanol were utilized for the pure synthesized sample, respectively. The absorbance at 515 nm was estimated using a 40 µL fraction of the sample-containing methanol
solution combined with 3 mL of the DPPH solution (Milton Roy, Spectronic 1201; Canada). The decrease in the absorbance at 515 nm was monitored continuously until it stabilized, with data
obtained at 1 min intervals for 16 min. The absorbance of the DPPH radical without an antioxidant was measured, as was the absorbance of ascorbic acid, a reference chemical. The percentage
inhibition (PI) of the DPPH radical was estimated using the Eq. (1): $${\text{PI}} = \left( {{\text{AC}} - {\text{AT}}} \right) \times {1}00$$ (1) where AC is the control absorbance at time
zero and AT is the sample absorbance plus DPPH at 16 min. The dose–response curve graphic plots were used to estimate the 50% inhibitory concentration (IC50), which is the concentration
necessary to inhibit the DPPH radical by 50%. The experiment was carried out in triplicate. COMPUTATIONAL METHODS The crystal structures of dihydrofolate reductase (PDB code: 5ISP32),
pyruvate kinase (PDB code: 5OE333), and sortase A (PDB code: 2MLM34) were obtained and utilized as templates for all _in-silico_ computations. To prepare these proteins, all inhibitors,
ions, and water molecules were removed. Modeller software was employed to construct all missing residues88. In addition, the H + + website was utilized to inspect the protonation states of
the studied targets89. All missing hydrogens were added. The structure of the investigated inhibitors was manually built. Prior to docking computations, all inhibitors were minimized using
the MMFF94S force field within SZYBKI software90,91. The atomic charges of the obtained compounds were determined utilizing the Gasteiger method92. All docking calculations were conducted
using AutoDock4.2.6 software93. Based on the AutoDock protocol, the investigated targets were saved in pdpqt format94. The number of genetic algorithm (GA) runs was adjusted to 250.
Moreover, energy evaluations (_eval_) were set to 25,000,000. The default settings of the other docking parameters were utilized. The grid box was set out to include the active site for all
investigated targets. STATISTICAL ANALYSIS The experimental data were recorded as an average value and SD, and each test was done in three replicates. The Duncan's multiple range test
was performed after the analysis of variance (ANOVA: two-factor with replication) and (t-Test: Two-Sample Assuming Equal Variances) on the data95,96. CONCLUSIONS In this study, _Aspergillus
pseudodeflectus_ AUMC 15761 used wheat bran in SSF to produce methyl ferulate and oleic acid for the first time, both of which have strong antibacterial and antioxidant properties. The most
prevalent chemical components were determined using gas chromatography-mass spectrometry (GC–MS) analysis of the crude methanol extract. An in-depth spectroscopic investigation was used to
identify methyl ferulate and oleic acid, which were then purified using column chromatography. Significant antioxidant and antibacterial properties of the two compounds against _Bacillus
subtilis_ and _Staphylococcus aureus_, respectively, were detected. These findings were ditto analyzed _in-silico_ utilizing the molecular docking technique. According to docking
computations, methyl ferulate, and oleic acid demonstrated good antibacterial activity. DATA AVAILABILITY All data related to this manuscript are incorporated in the manuscript only. CHANGE
HISTORY * _ 08 APRIL 2024 A Correction to this paper has been published: https://doi.org/10.1038/s41598-024-58336-9 _ REFERENCES * Li, D. _et al._ Ferulic acid: A review of its pharmacology,
pharmacokinetics and derivatives. _Life Sci._ 284, 119921 (2021). Article CAS PubMed Google Scholar * Lozano, R. _et al_. Global and regional mortality from 235 causes of death for 20
age groups in 1990 and 2010: A systematic analysis for the Global Burden of Disease Study 2010. _Lancet_ 380, 2095–2128 (2012). * Tang, Y., Hao, J., Fan, C. & Cao, X. Preparative
separation of high-purity trans-and cis-ferulic acid from wheat bran by pH-zone-refining counter-current chromatography. _J. Chromatogr. A_ 1636, 461772 (2021). Article CAS PubMed Google
Scholar * Malheiro, J. F., Maillard, J.-Y., Borges, F. & Simões, M. Evaluation of cinnamaldehyde and cinnamic acid derivatives in microbial growth control. _Int. Biodeter. Biodegrad._
141, 71–78 (2019). Article CAS Google Scholar * Pelaez, F. Biological activities of fungal metabolites. _Handb. Ind. Mycol._ 22, 41–92 (2004). Google Scholar * Newman, D. J. & Cragg,
G. M. Natural products as sources of new drugs from 1981 to 2014. _J. Nat. Prod._ 79, 629–661 (2016). Article CAS PubMed Google Scholar * Hawksworth, D., Lücking, R. Fungal diversity
revisited: 2.2 to 3.8 million species. _Microbiol. Spectr._ 5, FUNK-0052-2016 (2017). * Ismail, M. A., Moubasher, A. H., Mohamed, R. A. & Al-Bedak, O. A. Agro–industrial residues as
alternative sources for cellulases and xylanases production and purification of xylanase produced by _Aspergillus flavus_ AUMC 10331 isolated from extreme habitat. _Curr. Res. Environ. Appl.
Mycol._ 8, 313–322 (2018). Article Google Scholar * AL-Kolaibe, A. M., Moharram, A. M. & Al-Bedak, O. A. Worthwhile enzyme production and eco-friendly bioconversion of three
agricultural residues by _Aspergillus curvatus_ and _Aspergillus gaarensis_, promising enzyme-producers isolated from extreme environment. _J. Basic Appl. Mycol._ 12, 1–14 (2021). * Zhao, C.
H., Liu, X., Zhan, T. & He, J. Production of cellulase by _Trichoderma __reesei_ from pretreated straw and furfural residues. _RSC Adv._ 8, 36233–36238 (2018). Article ADS CAS PubMed
PubMed Central Google Scholar * Otari, S. V., Patel, S. K., Kalia, V. C. & Lee, J.-K. One-step hydrothermal synthesis of magnetic rice straw for effective lipase immobilization and
its application in esterification reaction. _Bioresour. Technol._ 302, 122887 (2020). Article CAS PubMed Google Scholar * Kothri, M. _et al_. Microbial sources of polyunsaturated fatty
acids (PUFAs) and the prospect of organic residues and wastes as growth media for PUFA-producing microorganisms. _FEMS Microbiol. Lett._ 367, fnaa028 (2020). * Fumagalli, F., Ottoboni, M.,
Pinotti, L. & Cheli, F. Integrated mycotoxin management system in the feed supply chain: Innovative approaches. _Toxins_ 13, 572 (2021). Article CAS PubMed PubMed Central Google
Scholar * Sultana, R. Ferulic acid ethyl ester as a potential therapy in neurodegenerative disorders. _Biochim. Biophys. Acta _1822, 748–752 (2012). * Phuong, N. T. M., Cuong, T. T. &
Quang, D. N. Anti-inflammatory activity of methyl ferulate isolated from _Stemona__ tuberosa_ Lour. _Asian Pac. J. Trop. Med._ 7, S327–S331 (2014). Article CAS Google Scholar * Pinheiro,
P. G. _et al._ Ferulic acid derivatives inhibiting _Staphylococcus aureus_ tetK and MsrA efflux pumps. _Biotechnol. Rep._ 34, e00717 (2022). Article CAS Google Scholar * D Archivio, M.
_et al_. Polyphenols, dietary sources and bioavailability. _Annali-Istituto Superiore di Sanita_ 43, 348 (2007). * Srinivasan, M., Sudheer, A. R. & Menon, V. P. Ferulic acid: Therapeutic
potential through its antioxidant property. _J. Clin. Biochem. Nutr._ 40, 92–100 (2007). Article CAS PubMed PubMed Central Google Scholar * Ou, S. & Kwok, K. C. Ferulic acid:
Pharmaceutical functions, preparation and applications in foods. _J. Sci. Food Agric._ 84, 1261–1269 (2004). Article CAS Google Scholar * Kikuzaki, H., Hisamoto, M., Hirose, K., Akiyama,
K. & Taniguchi, H. Antioxidant properties of ferulic acid and its related compounds. _J. Agric. Food Chem._ 50, 2161–2168 (2002). Article CAS PubMed Google Scholar * Moussouni, S.
_et al_. Crude peroxidase from onion solid waste as a tool for organic synthesis. Part II: Oxidative dimerization–cyclization of methyl p-coumarate, methyl caffeate and methyl ferulate.
_Tetrahedron Lett._ 52, 1165–1168 (2011). * Ghavam, M., Afzali, A. & Manca, M. L. Chemotype of damask rose with oleic acid (9 octadecenoic acid) and its antimicrobial effectiveness.
_Sci. Rep._ 11, 1–7 (2021). Article Google Scholar * Gochev, V. _et al_. Comparative evaluation of antimicrobial activity and composition of rose oils from various geographic origins, in
particular Bulgarian rose oil. _Nat. Prod. Commun._ 3, 1934578X0800300706 (2008). * Beltrán, G., Del Rio, C., Sánchez, S. & Martínez, L. Influence of harvest date and crop yield on the
fatty acid composition of virgin olive oils from cv. Picual. _J. Agric. Food Chem._ 52, 3434–3440 (2004). * Food, U., Administration, D. _FDA Completes Review of Qualified Health Claim
Petition for Oleic Acid and the Risk of Coronary Heart Disease_ (2018). * Barbour, J. A., Howe, P. R., Buckley, J. D., Bryan, J. & Coates, A. M. Effect of 12 weeks high oleic peanut
consumption on cardio-metabolic risk factors and body composition. _Nutrients_ 7, 7381–7398 (2015). Article CAS PubMed PubMed Central Google Scholar * Cater, N. B., Heller, H. J. &
Denke, M. A. Comparison of the effects of medium-chain triacylglycerols, palm oil, and high oleic acid sunflower oil on plasma triacylglycerol fatty acids and lipid and lipoprotein
concentrations in humans. _Am. J. Clin. Nutr._ 65, 41–45 (1997). Article CAS PubMed Google Scholar * Kim, J.-Y., Lee, H. W., Lee, S. M., Jae, J. & Park, Y.-K. Overview of the recent
advances in lignocellulose liquefaction for producing biofuels, bio-based materials and chemicals. _Bioresour. Technol._ 279, 373–384 (2019). Article CAS PubMed Google Scholar *
Mussatto, S. I., Ballesteros, L. F., Martins, S. & Teixeira, J. A. Use of agro-industrial wastes in solid-state fermentation processes. _Industrial Waste_ 274 (2012). * Praveen, V. &
Savitha, J. Solid state fermentation: an effective method for lovastatin production by fungi over submerged fermentation. _E3 J. Biotechnol. Pharm. Res. _3, 15–21 (2012). * Javed, S. _et
al._ Biotransformation of agricultural wastes into lovastatin and optimization of a fermentation process using response surface methodology (RSM). _Agronomy_ 12, 2848 (2022). Article CAS
Google Scholar * Reeve, S. M. _et al_. Charged propargyl-linked antifolates reveal mechanisms of antifolate resistance and inhibit trimethoprim-resistant MRSA strains possessing clinically
relevant mutations. _J. Med. Chem._ 59, 6493–6500. https://doi.org/10.1021/acs.jmedchem.6b00688 (2016). * Witzgall, F., Ewert, W. & Blankenfeldt, W. Structures of the N-terminal domain
of PqsA in complex with anthraniloyl- and 6-fluoroanthraniloyl-AMP: Substrate activation in _Pseudomonas_ quinolone signal (PQS). _Biosynthesis_ 18, 2045–2055, (2017). * Zhulenkovs, D.,
Rudevica, Z., Jaudzems, K., Turks, M. & Leonchiks, A. Discovery and structure–activity relationship studies of irreversible benzisothiazolinone-based inhibitors against _Staphylococcus
aureus_ sortase A transpeptidase. _Bioorg. Med. Chem._ 22, 5988–6003 (2014). Article CAS PubMed Google Scholar * Keller, N. P. Fungal secondary metabolism: regulation, function and drug
discovery. _Nat. Rev. Microbiol._ 17, 167–180 (2019). Article CAS PubMed PubMed Central Google Scholar * Evidente, A. _et al._ Fungal metabolites with anticancer activity. _Nat. Prod.
Rep._ 31, 617–627 (2014). Article CAS PubMed Google Scholar * Cole, J., Jarvis, B. & Schweikert, M. _Secondary Fungal Metabolites_ (Academic Press, 2003). Google Scholar * Sulyok,
M., Stadler, D., Steiner, D. & Krska, R. Validation of an LC-MS/MS-based dilute-and-shoot approach for the quantification of> 500 mycotoxins and other secondary metabolites in food
crops: Challenges and solutions. _Anal. Bioanal. Chem._ 412, 2607–2620 (2020). Article CAS PubMed PubMed Central Google Scholar * Thuan, N. N. _et al._ Secondary metabolites from the
fruiting bodies of _Coriolopsis__ aspera_ in Vietnam and their bioactivities. _Chem. Nat. Compd._ 57, 1104–1106 (2021). Article CAS Google Scholar * Woo, K. & Lee, K. CAS: 528: DC%
2BC3sXhslSmsLbP. _Nat. Prod. Sci_ 19, 221 (2013). CAS Google Scholar * Kwon, Y. S. & Kim, C. M. Antioxidant constituents from the stem of _Sorghum __bicolor_. _Arch. Pharm. Res._ 26,
535–539 (2003). Article CAS PubMed Google Scholar * Ilmiawati, A., Anggraini, D., Syahbirin, G., Rahayu, D. & Sugita, P. In _AIP Conference Proceedings._ 030009 (AIP Publishing LLC).
* Kohno, J. _et al._ Structures of TMC-120A, B and C, novel isoquinoline alkaloids from _Aspergillus __ustus_ TC 1118. _Tetrahedron_ 55, 11247–11252 (1999). Article CAS Google Scholar *
Kozlovskii, A. _et al._ Secondary metabolites of fungi of the Usti section, genus _Aspergillus_ and their application in chemosystematics. _Microbiology_ 86, 176–182 (2017). Article CAS
Google Scholar * Ogawa, A. _et al._ Pseudodeflectusin, a novel isochroman derivative from _Aspergillus __pseudodeflectus_ a parasite of the sea weed, _Sargassum fusiform_, as a selective
human cancer cytotoxin. _Bioorg. Med. Chem. Lett._ 14, 3539–3543 (2004). Article CAS PubMed Google Scholar * Rodrigues de Carvalho, C. _et al_. Biological activities of ophiobolin K and
6-epi-ophiobolin K produced by the endophytic fungus _Aspergillus calidoustus_. _Nat. Prod. Res._ 30, 478–481 (2016). * Asther, M. _et al._ Feruloyl esterase from _Aspergillus __niger_: A
comparison of the production in solid state and submerged fermentation. _Process Biochem._ 38, 685–691 (2002). Article CAS Google Scholar * Gaynes, R. The discovery of penicillin: New
insights after more than 75 years of clinical use. _Emerg. Infect. Dis._ 23, 849 (2017). Article PubMed Central Google Scholar * Bo, G. Giuseppe Brotzu and the discovery of
cephalosporins. _Clin. Microbiol. Infect._ 6, 6–8 (2000). * Kumar, C. G., Kamle, A., Mongolla, P. & Joseph, J. Parametric optimization of feruloyl esterase production from _Aspergillus
__terreus_ strain GA2 isolated from tropical agro-ecosystems cultivating sweet sorghum. _J. Microbiol. Biotechnol._ 21, 947–953 (2011). Article CAS PubMed Google Scholar * Keller, N. P.,
Turner, G. & Bennett, J. W. Fungal secondary metabolism—from biochemistry to genomics. _Nat. Rev. Microbiol._ 3, 937–947 (2005). Article CAS PubMed Google Scholar * Yang, X. _et
al._ Cyclosporine biosynthesis in _Tolypocladium_ _inflatum_ benefits fungal adaptation to the environment. _MBio_ 9, e01211-01218 (2018). Article CAS PubMed PubMed Central Google
Scholar * Brookman, J. L. & Denning, D. W. Molecular genetics in _Aspergillus fumigatus_. _Curr. Opin. Microbiol._ 3, 468–474 (2000). Article CAS PubMed Google Scholar * Talukdar,
R., Wary, S., Mili, C., Roy, S. & Tayung, K. Antimicrobial secondary metabolites obtained from endophytic fungi inhabiting healthy leaf tissues of _Houttuynia cordata_ Thunb., an
ethnomedicinal plant of Northeast India. _J. Appl. Pharm. Sci._ 10, 099–106 (2020). * Sengupta, S., Nandi, I., Bhattacharyya, D. & Ghosh, M. Anti-oxidant and anti-bacterial properties of
1-Octacosanol isolated from rice Bran Wax. _J. Plant Biochem. Physiol._ 6, 206 (2018). * Marik, T. _et al._ Structural diversity and bioactivities of peptaibol compounds from the
_Longibrachiatum__ clade_ of the filamentous fungal genus _Trichoderma_. _Front. Microbiol._ 10, 1434 (2019). Article PubMed PubMed Central Google Scholar * Nyfeler, R. &
Keller-Schierlein, W. Metabolites of microorganisms Echinocandin B, a novel polypeptide-antibiotic from _Aspergillus nidulans_ var. echinulatus: isolation and structural components.
_Helvetica Chim. Acta_ 57, 2459–2477 (1974). * Kalyani, P., Botsa, S. M., Laxmi, K. D. & Anil, S. Optimization of cultural conditions for biomass and antibacterial metabolite production
by _Aspergillus fumigatus_ strain MF1. _Hybrid Adv._ 2, 100016 (2023). * El-Sayed, A. S. _et al._ Production and bioprocess optimization of antitumor Epothilone B analogue from _Aspergillus
fumigatus_, endophyte of _Catharanthus roseus_, with response surface methodology. _Enzyme Microb. Technol._ 143, 109718 (2021). Article CAS PubMed Google Scholar * Ghavam, M.,
Azarnivand, H. & Akhbari, M. Examining of the quality and quantity of active ingredients of _Smirnova __iranica_ Sabeti in different habitats. _Health Biotechnol. Biophar_ 1, 21–27
(2018). Google Scholar * Rivaldi, J. D., Carvalho, A. K. F., da Conceição, L. R. V. & de Castro, H. F. Assessing the potential of fatty acids produced by filamentous fungi as feedstock
for biodiesel production. _Prep. Biochem. Biotechnol._ 47, 970–976 (2017). Article CAS PubMed Google Scholar * Thevenieau, F. & Nicaud, J.-M. Microorganisms as sources of oils. _Ocl_
20, D603 (2013). Article Google Scholar * Li, T. _et al._ Antibacterial properties of coaxial spinning membrane of methyl ferulate/zein and its preservation effect on sea bass. _Foods_
10, 2385 (2021). Article CAS PubMed PubMed Central Google Scholar * Jirovetz, L. _et al._ Chemical composition, antimicrobial activities and odor descriptions of some essential oils
with characteristic floral-rosy scent and of their principal aroma compounds. _Recent Res. Dev. Agron. Hortic._ 2, 1–12 (2006). CAS Google Scholar * Merkl, R., Hrádková, I., Filip, V.
& Šmidrkal, J. Antimicrobial and antioxidant properties of phenolic acids alkyl esters. _Czech J. Food Sci._ 28, 275–279 (2010). * Dufour, M. _et al._ Characterization of monolaurin
resistance in _Enterococcus faecalis_. _Appl. Environ. Microbiol._ 73, 5507–5515 (2007). Article ADS CAS PubMed PubMed Central Google Scholar * Speert, D. P., Wannamaker, L. W., Gray,
E. D. & Clawson, C. C. Bactericidal effect of oleic acid on group A streptococci: Mechanism of action. _Infect. Immunity_ 26, 1202–1210 (1979). Article CAS Google Scholar * Karamac,
M., Bucinski, A., Pegg, R. B. & Amarowicz, R. Antioxidant and antiradical activity of ferulates. _Czech J. Food Sci.-UZPI (Czech Republic)_ (2002). * Reddy, K. V. K. & Naidu, K. A.
Oleic acid, hydroxytyrosol and n-3 fatty acids collectively modulate colitis through reduction of oxidative stress and IL-8 synthesis; in _vitro_ and in _vivo_ studies. _Int.
Immunopharmacol._ 35, 29–42 (2016). Article CAS PubMed Google Scholar * Warcup, J. The soil-plate method for isolation of fungi from soil. _Nature_ 166, 117–118 (1950). Article ADS CAS
PubMed Google Scholar * Choi, Y.-W., Hyde, K. D. & Ho, W. Single spore isolation of fungi. _Fungal diversity_ (1999). * Smith, D. & Onions, A. H. _The Preservation and
Maintenance of Living Fungi_ (CAB International, 1994). * Houbraken, J. _et al._ Polyphasic taxonomy of _Aspergillus_ section Usti. _Stud. Mycol._ 59, 107–128 (2007). Article CAS PubMed
PubMed Central Google Scholar * Moubasher, A. H., Ismail, M. A., Al-Bedak, O. A. & Mohamed, R. A. _Ramophialophora_ _chlamydospora_, a new species from an alkaline lake of
Wadi-El-Natron, Egypt. _Asian J. Mycol._ 2, 110–117 (2019). Article Google Scholar * Al-Bedak, O. A. & Moubasher, A. H. _Aspergillus __gaarensis_, a new addition to section Circumdati
from soil of Lake El-Gaar in Wadi-El-Natron, Egypt. _Stud. Fungi_ 5, 59–65 (2020). Article Google Scholar * Katoh, K. & Standley, D. M. MAFFT multiple sequence alignment software
version 7: improvements in performance and usability. _Mol. Biol. Evol._ 30, 772–780 (2013). Article CAS PubMed PubMed Central Google Scholar * Criscuolo, A. & Gribaldo, S. BMGE
(Block Mapping and Gathering with Entropy): A new software for selection of phylogenetic informative regions from multiple sequence alignments. _BMC Evol. Biol._ 10, 210 (2010). Article
PubMed PubMed Central Google Scholar * Kumar, S., Stecher, G., Li, M., Knyaz, C. & Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. _Mol. Biol.
Evol._ 35, 1547–1549 (2018). Article CAS PubMed PubMed Central Google Scholar * Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. _Evolution_ 39,
783–791 (1985). Article PubMed Google Scholar * Posada, D. & Crandall, K. A. Modeltest: Testing the model of DNA substitution. _Bioinformatics_ 14, 817–818 (1998). Article CAS
PubMed Google Scholar * Al-Bedak, O. A. _et al._ Impact of fumigation with phosphine on viability of wheat grains stored for six months at two levels of moisture content, in addition to
description of four new records of associated fungi and assessment of their potential for enzymatic production. _J. Basic Appl. Mycol._ 11, 77–97 (2020). Google Scholar * Ramadan, A. M. _et
al._ GC-MS analysis, antifungal, and antioxidant activity of rice straw-based extract produced by _Aspergillus __tubingensis_ AUMC 15759 in solid state fermentation. _Egypt. Acad. J. Biol.
Sci. G_ 15, 43–60 (2023). Google Scholar * Azeem, M. _et al._ Development of one pot strategy for hyper production and in _vivo_ evaluation of lovastatin. _Molecules_ 25, 4380 (2020).
Article CAS PubMed PubMed Central Google Scholar * Ramadan, A. M. _et al._ Exploitation of sugarcane bagasse and environmentally sustainable production, purification, characterization,
and application of lovastatin by _Aspergillus __terreus_ AUMC 15760 under solid-state conditions. _Molecules_ 28, 4048 (2023). Article CAS PubMed PubMed Central Google Scholar * Chaman,
S., Sharma, G. & Reshi, A. K. Study of antimicrobial properties of _Catharanthus roseus_ by agar well diffusion method. _Int. Res. J. Pharm. Appl. Sci._ 3, 65–68 (2013). Google Scholar
* Siddiqui, B. S., Gulzar, T., Begum, S., Afshan, F. & Sattar, F. A. Insecticidal amides from fruits of _Piper nigrum_ Linn. _Nat. Prod. Res._ 19, 143–150 (2005). Article CAS PubMed
Google Scholar * Yen, G. C. & Duh, P. D. Scavenging effect of methanolic extracts of _peanut hulls_ on free-radical and active-oxygen species. _J. Agric. Food Chem._ 42, 629–632
(1994). Article CAS Google Scholar * Martí-Renom, M. A. _et al._ Comparative protein structure modeling of genes and genomes. _Annu. Rev. Biophys. Biomol. Struct._ 29, 291–325 (2000).
Article PubMed Google Scholar * Gordon, J. C. _et al._ H++: A server for estimating pKas and adding missing hydrogens to macromolecules. _Nucleic Acids Res._ 33, W368–W371.
https://doi.org/10.1093/nar/gki464 (2005). Article CAS PubMed PubMed Central Google Scholar * Halgren, T. A. M. M. F. F. V. I. MMFF94s option for energy minimization studies. _J.
Comput. Chem._ 20, 720–729 (1999). Article CAS PubMed Google Scholar * SZYBKI 1.9.0.3 (OpenEye Scientific Software, 2016). * Gasteiger, J. & Marsili, M. Iterative partial
equalization of orbital electronegativity: A rapid access to atomic charges. _Tetrahedron_ 36, 3219–3228. https://doi.org/10.1016/0040-4020(80)80168-2 (1980). Article CAS Google Scholar *
Huey, R. & Morris, G. M. _Using AutoDock 4 with AutoDocktools: a tutorial_ 54–56 (The Scripps Research Institute, 2008). Google Scholar * Forli, S. _et al._ Computational
protein-ligand docking and virtual drug screening with the AutoDock suite. _Nat. Protoc._ 11, 905–919. https://doi.org/10.1038/nprot.2016.051 (2016). Article CAS PubMed PubMed Central
Google Scholar * St, L. & Wold, S. Analysis of variance (ANOVA). _Chemom. Intell. Lab. Syst._ 6, 259–272 (1989). Article Google Scholar * Arthur, J. T. _Test Two Sample Assuming Equal
Variances in Excel_ (2009). Download references ACKNOWLEDGEMENTS Prof. Dr. M. T. Said is greatly appreciated for his kind help in all statistical analyses. FUNDING Open access funding is
provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB). This research was also funded by Researchers Supporting
Project number (RSP2024R364), King Saud University, Riyadh, Saudi Arabia. AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * Department of Botany & Microbiology, Faculty of Science, Al Azhar
University, Cairo, Egypt Ahmed Mohamed Ahmed Ali Ramadan, Reda Mohamed Shehata & Hussein Hosny EL-Sheikh * Department of Pharmacognosy, Faculty of Pharmacy, Al-Azhar University, Assiut
Branch, Assiut, 71524, Egypt Sabry Ahmed Hussein Zidan * The Regional Center for Mycology and Biotechnology (RCMB), Al Azhar University, Cairo, Egypt Reda Mohamed Shehata & Hussein Hosny
EL-Sheikh * Department of Botany & Microbiology, College of Science, King Saud University, 11451, Riyadh, Saudi Arabia Fuad Ameen * Department of Biological Sciences, University of
Arkansas, Fayetteville, USA Steven L. Stephenson * Assiut University Mycological Centre, Assiut University, Assiut, 71511, Egypt Osama Abdel-Hafeez Mohamed Al-Bedak Authors * Ahmed Mohamed
Ahmed Ali Ramadan View author publications You can also search for this author inPubMed Google Scholar * Sabry Ahmed Hussein Zidan View author publications You can also search for this
author inPubMed Google Scholar * Reda Mohamed Shehata View author publications You can also search for this author inPubMed Google Scholar * Hussein Hosny EL-Sheikh View author publications
You can also search for this author inPubMed Google Scholar * Fuad Ameen View author publications You can also search for this author inPubMed Google Scholar * Steven L. Stephenson View
author publications You can also search for this author inPubMed Google Scholar * Osama Abdel-Hafeez Mohamed Al-Bedak View author publications You can also search for this author inPubMed
Google Scholar CONTRIBUTIONS All authors participated equally to data analysis, authoring, and revising the article. The final version of the manuscript has been reviewed and approved by all
authors. CORRESPONDING AUTHORS Correspondence to Fuad Ameen or Osama Abdel-Hafeez Mohamed Al-Bedak. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests.
ADDITIONAL INFORMATION PUBLISHER'S NOTE Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. The original online
version of this Article was revised: The Funding section in the original version of this Article was incomplete. It now reads "Open access funding provided by The Science, Technology
& Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB). This research was also funded by Researchers Supporting Project number (RSP2024R364), King
Saud University, Riyadh, Saudi Arabia.”. RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use,
sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative
Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated
otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or
exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. Reprints
and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Ramadan, A.M.A.A., Zidan, S.A.H., Shehata, R.M. _et al._ Antioxidant, antibacterial, and molecular docking of methyl ferulate and oleic
acid produced by _Aspergillus pseudodeflectus_ AUMC 15761 utilizing wheat bran. _Sci Rep_ 14, 3183 (2024). https://doi.org/10.1038/s41598-024-52045-z Download citation * Received: 31 July
2023 * Accepted: 12 January 2024 * Published: 07 February 2024 * DOI: https://doi.org/10.1038/s41598-024-52045-z SHARE THIS ARTICLE Anyone you share the following link with will be able to
read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing
initiative






