- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT Cilia are protrusions of the cell surface and composed of hundreds of proteins many of which are evolutionary and functionally well conserved. In cells assembling motile cilia the
expression of numerous ciliary components is under the control of the transcription factor FOXJ1. Here, we analyse the evolutionary conserved FOXJ1 target CFAP161 in _Xenopus_ and mouse. In
both species _Cfap161_ expression correlates with the presence of motile cilia and depends on FOXJ1. Tagged CFAP161 localises to the basal bodies of multiciliated cells of the _Xenopus_
larval epidermis, and in mice CFAP161 protein localises to the axoneme. Surprisingly, disruption of the _Cfap161_ gene in both species did not lead to motile cilia-related phenotypes, which
contrasts with the conserved expression in cells carrying motile cilia and high sequence conservation. In mice mutation of _Cfap161_ stabilised the mutant mRNA making genetic compensation
triggered by mRNA decay unlikely. However, genes related to microtubules and cilia, microtubule motor activity and inner dyneins were dysregulated, which might buffer the _Cfap161_ mutation.
SIMILAR CONTENT BEING VIEWED BY OTHERS PROLIFERATION ASSOCIATED 2G4 IS REQUIRED FOR THE CILIATION OF VERTEBRATE MOTILE CILIA Article Open access 04 November 2024 FOXA1 IS A TRANSCRIPTIONAL
ACTIVATOR OF _ODF2/CENEXIN_ AND REGULATES PRIMARY CILIATION Article Open access 12 December 2022 PRIMARY CILIA AS DYNAMIC AND DIVERSE SIGNALLING HUBS IN DEVELOPMENT AND DISEASE Article 18
April 2023 INTRODUCTION Cilia are extensions that protrude from the cell surface of most vertebrate and some invertebrate cell types into the extracellular space. They can be broadly grouped
into nonmotile and motile cilia1. Nonmotile cilia, also referred to as primary cilia, are present on virtually every cell of vertebrates and are critical for signal transduction and sensing
of external stimuli (reviewed in2,3). Motile cilia move extracellular fluids along epithelia, or propel cells through the surrounding medium (e.g.4,5,6). Rotational movement of single
motile cilia on cells of the left–right organiser of vertebrate embryos generates a leftward fluid flow, which is pivotal for establishing the asymmetric arrangement of visceral organs7,8,9.
Coordinated whip-like beating of multiple cilia on ependymal cells of the brain, on epithelial cells of the respiratory tract, or of the fallopian tube are essential for cerebrospinal fluid
flow6,10,11,12, mucociliary clearance5,13, and movement of eggs into the ampulla and towards the uterus14, respectively. Wave-like movement of the sperm flagellum, a specialised long
cilium, is essential for sperm motility and fertilisation4. Defects in the formation of cilia or their function cause human diseases collectively known as ciliopathies (reviewed
in15,16,17,18,19). A specific subgroup of ciliopathies, referred to as primary cilia dyskinesia (PCD), is caused by impaired function of motile cilia. Reflecting the functions of motile
cilia, PCD is characterised by situs randomisation, impaired mucociliary clearance, respiratory problems, and male infertility. Mouse models of PCD frequently show reduced female fertility
and hydrocephalus12, which are less common in human patients (reviewed in20,21). The formation of apparently all motile cilia in the mouse as well as in other vertebrates and invertebrates
is under the control of the transcription factor FOXJ122,23,24,25,26,27,28. Thus, genes that act downstream of FOXJ1 are likely to be required for the formation or function of motile cilia.
In microarray screens for FOXJ1 target genes in node-stage embryos and foetal lungs29 we have identified a number of new _Foxj1_ targets30,31,32,33,34 including _Cfap161_, the orthologue of
the axonemal _Chlamydomonas reinhardtii_ FAP161 protein35. Recently, single-particle cryo-electron microscopy of _Chlamydomonas_ flagella showed that FAP161 is one of the 33 microtubule
inner proteins (MIPs) and likely localises to the A-tubule of the outer doublets36. Morpholino-mediated knock down of _Cfap161_ in zebrafish led to the loss of outer dynein arms, reduced
beating frequency of pronephric cilia and strong ciliopathy phenotypes including left–right asymmetry defects in one study37, but only a curved body axis and hydrocephalus in a subsequent
analysis38 leaving some uncertainties concerning the ciliary function of CFAP161. Human _CFAP161_ (c15orf26) is located on chromosome 15q in the linkage region of Kartagener syndrome39,
suggesting a potential involvement of _Cfap161_ mutations in this subtype of PCD. Here, we describe the analysis of CFAP161 function in _Xenopus_ crispants and mutant mice generated by
homologous recombination. Surprisingly, disruption of the highly conserved _Cfap161_ gene did not lead to obvious phenotypes related to dysfunctional motile cilia. RESULTS EXPRESSION OF
_CFAP161_ IN MOUSE AND _XENOPUS_ Mouse _Cfap161_ (RefSeq NM_029335.3) encodes an evolutionary conserved (Table S1) 303 amino acids (aa) protein lacking domains of known biochemical function.
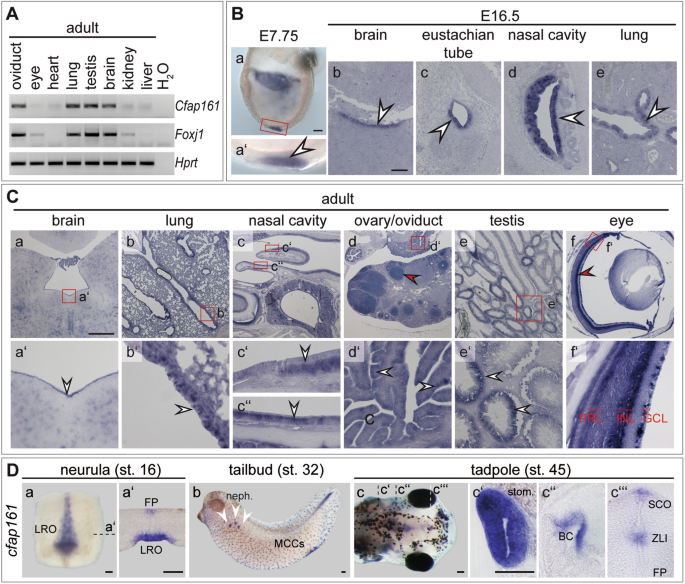
High expression levels of _Cfap161_ correlate in general with the presence of motile cilia and co-expression of _Foxj1_ (Fig. 1A). In early embryos (E7.75) _Cfap161_ is expressed in the
ciliated ventral layer of the embryonic node (the mouse left–right organiser, LRO; Fig. 1Ba,a′), and in later foetal stages (E16.5) it is expressed in ependymal cells of the brain (Fig.
1Bb), epithelial cells lining the eustachian tube (Fig. 1Bc), the respiratory epithelium of the nasal cavity (Fig. 1Bd), and in ciliated epithelial cells of the lung (Fig. 1Be). In these
tissues expression was also detected in adults (Fig. 1A) as well as in cells of the oviduct carrying motile cilia, and in testis (Fig. 1A,Ca–e). In testis, _Cfap161_ transcripts were not
found in all seminiferous tubules (Fig. 1Ce,e′) suggesting that _Cfap161_ transcription is restricted to distinct stages of the cycle of the epithelium of the seminiferous tubules.
Consistent with this idea, CFAP161 protein was only observed at late stages of spermiogenesis (Fig. S1). _Cfap161_ mRNA was additionally detected in some cells lacking motile cilia: in the
retina, in the ganglion cell layer (GCL), the inner nuclear layer (INL), in the photoreceptor cells (PRL; Fig. 1Cf, red arrowhead in f′) and in developing follicles (red arrowhead in Fig.
1Cd). Likewise, CFAP161 protein was detected in cells harbouring immotile cilia (primary cilia), such as hair cells in the inner ear or in cells of the kidney collecting ducts (Fig. S2).
Consistent with the prominent expression in cells carrying motile cilia in mouse tissues, _cfap161_ was detected in the left–right organiser (LRO; Fig. 1Da,a′) and in the floor plate of the
prospective neural tube (Fig. 1Da′) during neurulation in _Xenopus laevis_ embryos. Tailbud stages showed transcripts in multiciliated cells (MCCs) of the epidermis and in the nephrostomes
(Fig. 1Db). Free-swimming tadpoles established expression domains in the stomach (stom.; Fig. 1Dc,c′), branchial chamber (BC; Fig. 1Dc,c″), subcommissural organ (SCO) and in the zona
limitans intrathalamica (ZLI; Fig. 1Dc,c‴). LOCALISATION OF CFAP161 AND DEPENDENCE ON FOXJ1 To study CFAP161 protein expression in mouse we generated a monoclonal rat antibody directed
against a peptide (α-pI) mainly encoded by exon 1 (Fig. 2A and Fig. S3) and polyclonal antibodies directed against a peptide (α-pII) encoded by exon 2 in rabbits (Fig. 2A). In Western blots
both antibodies detected Flag-tagged CFAP161 over-expressed in CHO cells as well as endogenous CFAP161 in testis lysates (Fig. 2B). Additionally, these antibodies detected CFAP161 expression
concomitantly with cilia formation indicated by co-expression with the ciliary protein IFT88 in air–liquid interface (ALI) cultures of mouse tracheal epithelial cells (mTECs) (Fig. 2C). The
monoclonal antibody detected endogenous CFAP161 in foetal (E17.5) tissue sections (Fig. 2D), consistent with _Cfap161_ mRNA expression domains (Fig. 1Bb–e) corroborating the specific
expression of _Cfap161_ in cells carrying motile cilia. CFAP161 was not detected in tissues of _Foxj1_ null mutant (_Foxj1__lacZ/lacZ_) foetuses (n = 4; Fig. 2De–h). Likewise, _Xenopus
foxj1_-crispants revealed a global downregulation of _cfap161_ as prominently seen in the epidermis (Fig. 2Eb,b′) where expression of _cfap161_ is nearly absent in the MCCs, and a unilateral
_foxj1_ gain of function (_foxj1_-GOF) strongly induced transcription of _cfap161_ (Fig. 2F) confirming the dependence of _Cfap161_ expression on FOXJ1 as was previously observed in
zebrafish37. Indirect immunofluorescence of adult tissue sections using α-pI and α-pII antibodies showed co-localisation of CFAP161 with acetylated-α-tubulin (ac-TUB; Fig. 3A), indicating
that CFAP161 is a component of motile cilia. CFAP161 and ac-TUB staining did not overlap in the distal part of cilia (Fig. 3Aa′–d′, arrowheads in insets) indicating that CFAP161 is excluded
from the tip of the cilium, which was most clearly seen in the sections of the nasal respiratory epithelium (Fig. 3Ac′,d′). Absence of CFAP161 from the cilium tip is consistent with the
localisation of its _Chlamydomonas_ orthologue in outer microtubule doublets and likely reflects the fact that outer doublets terminate earlier than the central inner pair40. Consistent with
the ciliary localisation, CFAP161 was found in the flagellum of spermatozoa (Fig. 3Ae,e′; Fig. S1). Detection of _Xenopus_ CFAP161 failed with α-pI and α-pII antibodies but heterologously
expressed murine CFAP161 tagged with a N-terminal EGFP (GFP-CFAP161) partially co-localised with a Cetn4-RFP fusion construct, which marks the basal bodies of epidermal MCCs (Fig. 3Ba). Of
note, most GFP-CFAP161 signal was in anterior juxtaposition underneath each basal body (Fig. 3Ba′,a″), reminiscent of the rootlet (Fig. 3Bb). Localisation at basal bodies might reflect
non-physiological accumulation due to high levels of the overexpressed GFP-tagged CFAP161. Similarly, also overexpressed N- or C-terminally HA-tagged _Xenopus_ Cfap161 was not specifically
localised in cilia but found throughout the whole cell (Fig. S4). It appears unlikely that in _Xenopus_ this conserved ciliary protein is absent from motile cilia. Rather, the apparent
uniform distribution of HA-tagged Cfap161 might reflect high levels of overexpressed HA-tagged Cfap161 that exceed the amount that can translocate to cilia. To obtain first insights into the
biochemical context in which CFAP161 might act, we identified potential interaction partners by mass spectrometry of CFAP161 complexes immunoprecipitated from wild type testis lysates using
the polyclonal α-pII antibody (Table S2). One potential interaction partner was KIAA0556 (also known as Katanin-interacting protein KATNIP), a basal body protein that stabilises cytoplasmic
microtubules in human cells, regulates ciliary A-tubule number in _C. elegans,_ and when mutated, causes Joubert syndrome in humans41. Yeast-two-hybrid analysis (Fig. S5) validated a robust
interaction of CFAP161 and KIAA0556, supporting a function of CFAP161 in some aspects of microtubule organisation or function. FUNCTIONAL ANALYSES OF CFAP161 IN _XENOPUS_ To investigate the
function of CFAP161 in the frog _Xenopus_ we mutated _cfap161_ by genome editing using CRISPR/Cas9. Two different single guide RNAs (sgRNAs) were designed, targeting exon 1 (sgRNA1) and
exon 3 (sgRNA2), respectively. Sequencing of PCR products with pooled DNAs of 10 F0 crispants (Fig. S6A,B) confirmed successful genome editing of _cfap161_. Furthermore, _cfap161_
transcripts were strongly reduced in _cfap161_-crispants, indicative for nonsense mediated mRNA decay (Fig. S6Cb,b′). Initial experiments with individual sgRNAs showed no effect in ciliated
cells of the larval epidermis (data not shown). Subsequently both sgRNAs (sgRNA1 & sgRNA2) were injected simultaneously. Crispants were analysed for laterality defects, the development
of pronephric cysts or an externally visible hydrocephalus in stage 45 tadpoles. Evaluation of 297 crispants obtained in five independent experiments showed no impact on their overall
phenotype (Fig. 4A–D). To address whether loss of _cfap161_ has a more subtle effect on ciliary beating of epidermal MCCs, we analysed high speed video microscopy recordings of stage 32 wild
type and crispant specimens for ciliary beat frequency (CBF) and cilia generated flow (CGF) as described in detail in33. Neither CBF (approximately 22 Hz in both wild type and crispant
embyos; Fig. 4Ea,b; Table S3) nor CGF (around 400 µm/s for both wild type and crispant embryos; Fig. 4Fa,b; Table S4) was affected by mutating _cfap161_. Thus, disruption of _cfap161_ showed
no phenotypic changes associated with dysfunctional motile cilia in _Xenopus_. FUNCTIONAL ANALYSIS OF CFAP161 IN MICE To analyse CFAP161 function under physiological conditions in mice we
generated a conditional allele by flanking _Cfap161_ exons 2 and 3 by loxP recombination sites (Fig. 5A). Deletion of these two exons by Cre-mediated site-specific recombination results in a
frame shift that generates a premature translational stop codon by the first base triplet of exon 4, which should effectively abolish the generation of full-length CFAP161 protein. To
generate mice lacking CFAP161 in all tissues we deleted exons 2 and 3 (_Cfap161__∆ex2,3_) in the female germ line using ZP3:Cre mice42. Homozygous _Cfap161__∆ex2,3_ mice were born at
Mendelian ratio (wt 95, het 186, hom 91; χ2 = 0.086, p = 0.98) and showed no obvious abnormalities. Western blot analyses of testis and epididymis lysates with the monoclonal antibody α-pI
(which recognises the epitope encoded by exon 1; analysis shown in Fig. S3) and with polyclonal antibody α-pII did not detect full length CFAP161 protein or any other shorter protein product
(Fig. 5B; Fig. S9C) that might have been generated by alternative splicing. Likewise, our antibodies did not detect CFAP161 protein in mutant testis sections (Fig. 5Cb,d) supporting that
deletion of exons 2 and 3 abolishes production of CFAP161 and thus leads to a functional null allele. Disruption of normal motile cilia function can lead to hydrocephalus and mucus
accumulation in the respiratory tract, abnormal _situs_ of visceral organs and male infertility due to immotile spermatozoa, rarely female infertility20. Homozygous mutants showed no
externally visible abnormalities over an observation period of ≥ 8 months, and matings of homozygous breeding pairs (n = 3) gave rise to litters with normal sizes. Serially sectioned brains
(section plane is shown in Fig. 6A) of 4 months old _Cfap161__∆ex2,3_ homozygotes (n = 4) did not show signs of enlarged ventricles (Fig. 6Cb,b′,d,d′,d″). Likewise, sections of mutant lungs
(n = 4; Fig. 6Cf,f′) or PAS stained serial sections (section planes are shown in Fig. 6B) of the mutant nasal cavities (Fig. 6Ch,h′,j) did not show obvious morphological alterations or
accumulation of mucus. Although mutant males bred apparently normal computer-assisted sperm analysis (CASA) was performed to detect potential abnormalities in sperm number or motility but
revealed no differences between wild type and mutant (Fig. S7A; Table S5) samples. Likewise, electron microscopic analyses of mutant sperm cells prepared from cauda epididymis showed no
obvious structural abnormalities (Fig. S7B) in the axonemes of flagella and cilia of the lung (Fig. S7C). Additionally, no polydactyly or deafness as a sign for impairment of primary cilia
were observed in the mutants (n = 31). Taken together, our mutational analyses in _Xenopus_ and mice did not reveal any evidence for essential CFAP161 functions in motile cilia in either
vertebrate species. RNA-SEQ ON CFAP161-DEFICIENT TESTIS REVEALS DYSREGULATION OF CILIARY GENE EXPRESSION Given the unexpected result that the mutation of an evolutionary highly conserved
FOXJ1 target gene has no motile cilia-related phenotype, we analysed transcriptional changes in control wild type and _Cfap161__∆ex2,3/∆ex2,3_ testes by mRNA-sequencing (RNA-seq).
Differential expression analysis using DEseq2 revealed a number of statistically significantly dysregulated transcripts (338; P-adj < 0.05) (Table S6). Gene-ontology (GO) analysis43,44 of
these transcripts showed a significant enrichment of biological process terms relating to microtubule and cilia, including mitotic spindle organisation, protein localisation to
cytoskeleton, microtubule bundle formation, cilium movement, and cytoskeleton-dependent intracellular transport (Table S7). Additionally, terms related to RNA biology (e.g. mRNA
stabilisation, regulation of RNA processing, regulation of RNA splicing) as well as to chromatin regulation (e.g. chromatin remodelling, DNA packaging, and histone modification) were
enriched. The most enriched GO-terms were ATP-dependent microtubule motor activity (minus-end-directed) and inner dynein arm when the dataset was compared to GO molecular function and GO
cellular component datasets, respectively. Of the 338 dysregulated genes, 164 were downregulated and 174 were upregulated, but only few transcripts showed strong changes in expression levels
(3 transcripts showed downregulation to levels of 0.5-fold-expression or smaller, and 6 showed upregulation of 2-fold-expression or higher, including _Cfap161_) (Table S8). DISCUSSION In
this analysis we show that strong _Cfap161_ expression correlates well with the presence of motile cilia, consistent with its dependence on FOXJ1 both in _Xenopus_ and mice. In mice CFAP161
protein localises to the axoneme, consistent with localisation of its _Chlamydomonas_ orthologue FAP161 as a microtubular inner protein of the A-tubule of outer doublet microtubules. In both
analysed vertebrate species, the disruption of _Cfap161_ did not result in phenotypes related to dysfunctional motile cilia. Comparison of mRNA from wild type and _Cfap161_-mutant mouse
testis showed stabilisation or enhanced transcription of _Cfap161_ mRNA in mutants and dysregulation of more than 300 genes including genes related to microtubules and cilia, microtubule
motor activity and inner dyneins. The absence of phenotypes related to dysfunctional motile cilia in both _Xenopus_ and mouse came as a surprise given that morpholino-mediated knock down of
_cfap161_ (previously referred to as _C18H15orf26)_ caused strong37 and mild38 ciliopathy phenotypes in zebrafish. Also, the interaction of CFAP161 with KIAA0556, a basal body and
microtubule-associated protein which stabilises cytoplasmic microtubules in human cells and regulates ciliary A-tubule number in _C. elegans_, supported a ciliary function of CFAP161,
although a truncation of the KIAA0556 protein was not associated with defective cilia structure or motility in ventricular ependymal cells in mice as well41. One potential explanation for
the lack of _Cfap161_ mutant phenotypes in _Xenopus_ and mouse could be the presence of redundant genes. However, we did not find any evidence for the presence of _Cfap161_ paralogs in both
species. To investigate the function of _cfap161_ in the frog _Xenopus_, we initially also used Morpholino oligomers (MOs) that interfere with either mRNA translation or splicing (TBMO or
SBMO). However, only high MO doses resulted repeatedly in specimens with enlarged brain ventricles, oedema and compromised ciliation of epidermal MCCs, and we were unable to rescue any of
these phenotypes with a tagged murine or untagged _Xenopus cfap161_ mRNA (data not shown). Because these phenotypes were not observed after disruption of _cfap161_ by CRISPR/Cas9 we
attribute these phenotypes to unspecific effects of the used morpholinos at high concentrations. A ciliary function of _cfap161_ in zebrafish that has been lost in _Xenopus_ and mice cannot
be ruled out at present, however it seems unlikely. Thus, it is possible that the discordant observations after morpholino knock down of _cfap161_ in zebrafish37,38 reflect unspecific
effects similar to those we observed in _Xenopus_. Genetic compensation of mutations is increasingly observed in various species (reviewed in45). _Xenopus cfap161_-crispants showed
downregulation of _cfap161_ mRNA (Fig. S6Cb,b′) suggesting nonsense-mediated decay. mRNA degradation is known to trigger genetic compensatory mechanisms in zebrafish embryos and mouse
cells46. Thus, in principle, the absence of a motile cilia related phenotype in _Xenopus_ crispants might be the result of genetic compensation triggered by _cfap161_ mRNA decay. However,
the absence of a specific effect in _cfap161_ morphants argues against this possibility. Also, our RNA-seq data from the mouse do not support the action of a compensatory mechanism triggered
by nonsense-mediated mRNA decay because _Cfap161_ mRNA was stabilised rather than destabilised in mutants. Thus, if genetic compensation underlies the absence of a phenotype in
_Cfap161_-mutant mice the mechanism cannot depend on mRNA degradation46. mRNAs bearing a premature termination codon as is the case in our _Cfap161__∆ex2,3_ allele can trigger genetic
compensation by promoting transcription of homologous genes47. However, as stated before, no _Cfap161_ paralogs were found in the mouse genome making this possibility unlikely. The high
number of statistically significantly dysregulated transcripts in _Cfap161_-mutant testis resulting from mutation of a gene without known transcriptional activity is surprising as is the
abundance of these dysregulated genes on chromosome 7, where also _Cfap161_ is located. Analysis of gene ontology and GO molecular function and GO cellular component datasets suggested a
potential effect of CFAP161 loss of function on the expression of other genes including genes related to microtubules and cilia, microtubule motor activity and inner dyneins. A conclusive
interpretation of the RNA-seq results seems difficult as to the precise mechanism of why expression of so many genes is affected and how that might affect the loss of _Cfap161_ phenotype and
potentially buffer the _Cfap161_ mutation. While protein complexes bound to the inner surface of doublet microtubules were identified in diverse species (reviewed in48) not much is known
about the biological functions of these proteins. In _Tetrahymena_ RIB72A and the conserved RIB72B are MIPs that bind to the inner surface of A-tubules. Deletion of either protein caused
disruption of distinct MIP complexes, severe structural defects of the A-tubule and abnormal ciliary beating indicating MIP functions in stabilisation of doublet microtubules and possibly
regulation of ciliary beating49. In contrast, single deletion of the conserved B-tubule MIPs FAP45 and FAP52 had a slight or no effect, respectively, on swimming velocity and ciliary beat
frequency in _Chlamydomonas_, whereas only deletion of both MIPs significantly reduced both parameters and destabilised B-microtubules, which was suggested to reflect a “fail-safe” mechanism
acting in the stabilisation of doublet microtubules50. Such a “fail-safe” mechanism could possibly also explain the apparent absence of a ciliary phenotype in _Cfap161_ mutants. We cannot
rule out that disruption of _Cfap161_ causes subtle defects of stability or function of motile cilia. Such defects, if present, might sensitise cilia for other mutations or only manifest
themselves under other conditions impinging on ciliary function. MATERIAL AND METHODS ETHICS APPROVAL Mouse and _Xenopus laevis_ handling and husbandry was in accordance with the German
regulations (Tierschutzgesetz) and for mice approved by the ethics committee of Lower Saxony for care and use of laboratory animals (LAVES, Niedersächsisches Landesamt für Verbraucherschutz
und Lebensmittelsicherheit), and for frogs by the Regional Government Stuttgart, Germany (A379/12 Zo ‘Molekulare Embryologie’). Mice were kept in the central animal facility of Hannover
Medical School (ZTL) as approved by the responsible Veterinary Officer of the City of Hannover. All experiments were performed in accordance with the relevant guidelines and regulations and
in compliance with the ARRIVE guidelines. STATISTICAL ANALYSES Statistical analyses were done using Prism7 for Student’s paired t-test. The chi-square or Wilcoxon-Match-Pair test was done in
RStudio (The R project for statistical computing; http://www.r-project.org/). The used tests are indicated in the respective figure legends. Statistical analysis for proteomics data was
done using Perseus. MOUSE METHODS EXPERIMENTAL ANIMALS _Foxj1-_mutant _(Foxj1__lacZ_)22, Zp3:Cre42, and FLPe mice51 were described previously. _Cfap161__loxP_ mice were generated by Cyagen
(Cyagen Biosciences, California, USA). The neo cassette was flanked by FRT sites and removed by FLP-mediated recombination. Exon 2 and 3 were deleted in the germline of _Cfap161__loxP_;
Zp3:Cre42 double heterozygous females. _Cfap161__∆ex2,3_ mice were analysed on a hybrid (CD-1//129/Sv) genetic background. MOUSE GENOTYPING _Cfap161_-mutant and wild type mice were genotyped
by PCR with primer pairs: * Cfap161-loxP-F1: 5′-ACCTTTGCTACCCGAGGTAGTTC-3′, Cfap161-loxP-R1: 5′-ATGCATAAGGAAGGGAAGGATAGG-3′, 312 bp wild type, and 372 bp _Cfap161__loxP_ product. *
Cfap161-loxP-F1: 5′-ACCTTTGCTACCCGAGGTAGT-3′, Cfap161Neo del-R1: 5′-CCTGCGGATCATTTCCAAAACCTC-3′, 515 bp _Cfap161__∆ex2,3_ product. COLLECTION, EMBEDDING, AND SECTIONING OF MOUSE TISSUES Mice
were killed by cervical dislocation, tissues dissected, fixed overnight at 4 °C in 4% PFA, 100% methanol or 100% methanol/DMSO (4:1), if necessary decalcified in 0.5 M EDTA for 2 weeks
exchanging EDTA every other day, dehydrated, embedded in paraffin according to standard procedures and sectioned at 5 or 10 µm. For standard histology the lungs were not inflated, but wt and
mutant were equally treated. Mice used for transmission electron microscopy (TEM) were perfused with fixative followed by dissection of the organs and embedded and analysed as described52.
See “Supplemental Material and Methods” for details. SECTION IN SITU HYBRIDISATIONS (SISH) SISH were performed on 10 µm sections of PFA fixed and paraffin-embedded tissues as described in53.
DIG-labelled RNA probe was produced from FANTOM plasmid ZX00121F1054 using the Roche DIG RNA labelling system. WHOLE MOUNT IN SITU HYBRIDISATION (WISH) WISH were performed on E7.5–8.0 old
mouse embryos as described29. HISTOLOGICAL STAINING 5 µm sections of PFA fixed and paraffin-embedded tissues were stained with haematoxylin and eosin (HE) according to standard procedures,
or with PAS using the Periodic Acid-Schiff Kit (Sigma Aldrich) according to manufacturer’s instructions. CELL CULTURE Murine tracheal epithelial cells (mTECs) were isolated and cultured at
air liquid interface (ALI) as described55, and cultured for up to 10 days. CHO cells were cultured in DMEM/F12 containing 10% FCS, Pen/Strep, and 2 mM Glutamax (Gibco). CHO cells were
transfected using Perfectin (Genlantis) according to manufacturer’s instructions. RNA ISOLATION AND RT-PCR Total RNA was isolated using Direct-zol RNA MiniPrep Kit (ZYMO) according to the
manufacturer’s instructions. cDNA was produced using the Superscript-II Reverse Transcriptase kit (Thermo Fisher Scientific). PCRs were performed using primer combinations: mouse _Hprt_
(exon 7–9; 249 bp) (5′-CACAGGACTAGAACACCTGC-3′; 5′-GCTGGTGAAAAGGACCTCT-3′), mouse _Cfap161_ (exon 4–6; 230 bp) (5′-ATAGCAGCAGGACTGGAAGGCAAG-3′; 5′-GCTCGGTTTGTGTGACGATGATAG-3′), mouse _Foxj1_
(exon 2–3; 432 bp) (5′-CTTCTGCTACTTCCGCCATGC-3′; 5′-TCCTCCTGGGTCAGCAGTAAGG-3′). _Hprt_-RT-PCR was used to validate the integrity of the RNA. RNA ISOLATION AND RNA-SEQ Total RNA was isolated
from single testis without epididymis from 3 different 11–17 weeks old wild type and 3 different 11–13 weeks _Cfap161__∆ex2,3/∆ex2,3_ males using the Direct-zol RNA MiniPrep Kit (ZYMO)
according to manufacturer’s instructions. RNA integrity was first validated by gel electrophoresis, then further analysed by Eukaryote Total RNA Nano and Bioanalyser. Details concerning
library generation, quality control and quantification as well as details for the sequencing run and raw data processing can be found in the “Supplementary Materials and Methods”. GENERATION
OF ANTIBODIES Monoclonal antibodies (MAbs) against mouse CFAP161 were generated as described in33 by immunisation of rats with the peptide RMGNWNEDVYLEEERM (pI; aa11-26). MAbs that reacted
specifically with CFAP161 were further analysed by Western blots and indirect immunofluorescence. α-CFAP161 pI clone 8F9 was further characterised (see Fig. S3 and “Supplemental Methods” for
details) and used in this study. Rabbit polyclonal antibodies against mouse CFAP161 epitope ELLIQRNRRVKKNIL (pII; aa37-51) and YLDSHKVEKPKNQW (pIII; aa254-267) in two different animals (a1
and a2) were generated and affinity purified by BioGenes (BioGenes GmbH, Berlin). IMMUNOFLUORESCENCE STAINING Sections (5 µm) were deparaffinised, rehydrated and antigens unmasked by boiling
for 20 min in 10 mM Tris–HCl pH 9.5, 1 mM EDTA. Unspecific binding was blocked by 5% FCS in PBS. Primary antibodies were incubated at 4 °C overnight, secondary antibodies for 1 h at room
temperature. Antibodies were diluted in blocking solution: α-acetylated α-tubulin (ac-TUB) (Sigma Aldrich T6793) 1:1000; α-CFAP161 pI 1:50; α-CFAP161 pII 1:2000; α-mouse-Alexa633 (Invitrogen
A21052) 1:500; α-mouse-Alexa555 (Life Technologies A31570) 1:500; α-mouse-Alexa555 (Invitrogen A21424) 1:500; α-rat-Alexa488 (Invitrogen A21208) 1:500; α-rabbit-Alexa488 (Invitrogen A11034)
1:500; and α-rabbit-Alexa555 (Invitrogen A21429) 1:500. DAPI (1 µg/ml, Applichem) and Lectin-PNA-Alexa488 (Thermo Fisher L-21409) 1:500 were incubated together with secondary antibodies.
WESTERN BLOT ANALYSIS SDS-PAGE and Western blotting were carried out according to standard procedures. The following amounts of total proteins were loaded per lane: CHO cells (Fig. 2B and
Fig. S9A) and testis or epididymis lysate (Figs. 2B, 5B, Fig. S9A,C) 15–20 µg. For mTEC ALI cultures (Fig. 2C and Fig. S9B) 1/10 of the lysate of a confluent 24-well transwell insert was
loaded, which amounts to 2.5–4 µg according to55. Blots were blocked with 5% milk in PBS/0.1% Tween20. Antibodies were diluted in blocking solution: α-β-TUBULIN (β-TUB; Sigma; T7816)
1:250,000, α-IFT88 (Proteintech; 13967-1-AP) 1:1000, α-CFAP161-pI 1:100, α-CFAP161-pII 1:2000, α-mouse-POD (Amersham NA931) 1:10,000; α-rabbit-POD (Amersham NA934) 1:10,000, and α-rat-POD
(Amersham NA935) 1:10,000. Western blots were developed using WesternBright Quantum (Advansta). YEAST TWO-HYBRID (Y2H) ASSAY TO VALIDATE CFAP161-KIAA0556 INTERACTION Yeast two-hybrid assays
were performed according to the manufacturer’s protocol (Matchmaker Gold Yeast Two-Hybrid System 630489, Clontech). For details see “Supplemental Methods”. IMMUNOPRECIPITATIONS (IPS) AND
MASS SPECTROMETRY (MS) CFAP161 was immunoprecipitated from wild type mouse testis with epididymis using the Thermo Scientific Pierce crosslink IP kit using α-CFAP161 pII or with rabbit-IgG
as controls. In each experiment (n = 3) the total lysate of one homogenised (160–200 mg) testis and epididymis (~ 8–10 mg total protein) was incubated either with IgG or affinity-purified
α-CFAP161 pII crosslinked to resin. 1 mg of protein was loaded to 40 µl settled resin as suggested by the manufacturer’s instructions (in practice, 2 mg of total protein was loaded per
column containing 80 µl of resin) in n = 6 IgG control IPs and n = 7 CFAP161 IPs. Immunoprecipitated CFAP161 complexes were isolated, purified, and subsequently analysed by LC–MS/MS as
described34. _XENOPUS_ METHODS WHOLE MOUNT IN SITU HYBRIDISATION (WISH) WISH were performed on MEMFA fixed embryos covering the first week of development. DIG-labelled RNA probe was produced
from I.M.A.G.E. clone plasmid IRBHp990H1063D54 using the Roche DIG RNA labelling system. GENERATION AND VERIFICATION OF CRISPANTS _cfap161_ sgRNAs targeting exon 1 and exon 3 were designed
using CRISPRscan. DNA templates were created using the Promega Pfu proofreading polymerase for oligo extension reaction. sgRNA synthesis utilised the Invitrogen mMESSAGE mMACHINE T7
Transcription Kit in combination with the Invitrogen MEGAclear Transcription Clean-Up Kit. 300 ng _foxj1_33 or 150 ng of each _cfap161_ sgRNAs were preassembled with 1 ng PNABio Cas9 protein
from _Streptococcus pyogenes_ with NL and injected at the 1 cell stage. Genome editing was confirmed via Synthego ICE analysis (https://ice.synthego.com) after direct sequencing of PCR
products from pooled stage 45 embryos using the following primer combinations: _cfap161_ target site exon 1 (5′-CGTCGCCTTGGCAACTGATA-3′; 5′-TGGGGGTCGGATTTTCAATG-3′), _cfap161_ target site
exon 3 (5′-TTTGGGGATGTGGTGATGCT-3′; 5′-CGTCACACTCAGTGACATCAT-3′). CFAP161/CFAP161 LOCALISATION 20 pg of either pCS2 + EmGFP-_Cfap161,_ pCS2 + HA-_cfap161 or_ pCS2 + _cfap161_-HA were
co-injected with 20 pg pCS2 + _cetn4_-RFP or pCS2 + GFP-_cetn4_ at the 4 cell stage into the ventral marginal zone to target the epidermis. Embryos were fixed in MEMFA and stained with
Thermo Fisher Scientific Alexa Fluor 405 Phalloidin to mark the F-actin of the cell borders before fluorescence imaging. CFAP161- and CETN4-localisation was shown by direct fluorescence.
HIGH-SPEED VIDEO MICROSCOPY OF LARVAL EPIDERMAL CILIA Videos were recorded at stage 32 embryos using a Zeiss Axioskop 2 mot plus microscope. Embryos were mounted on a slide containing a
chamber constructed from duct tape. The most ventral part of the belly was recorded with a high-speed Hamamatsu video camera Orca flash 4.0 at 800 frames per second (fps) for 1 s to analyse
ciliary beating. Ciliary flow was analysed using 1 μm fluorescent beads (Invitrogen FluoSpheres; 1:2000). Beads were added to the culture medium (0.1 × MBSH) and specimens were imaged using
a Zeiss Axiocam HSm camera at 175 fps. Evaluation of CBF and bead transport was previously reported in33. DOCUMENTATION WISH, SISH, histological stainings and IHC on sections of all mouse
experiments were documented with a Leica DM5000 microscope equipped with a Leica DFC300FX digital camera and the used software was Leica FireCam (Version 1.9.1). Immunofluorescence staining
of mouse tissue sections were photographed with the Leica DMI6000 microscope equipped with a Leica DFC350FXR2 camera and the Leica Application Suite X software. Images were processed in
Adobe Photoshop CS4 or CC, or FIJI ImageJ version 2.0.0, figures assembled using Adobe Illustrator CS4, CS6 or CC. _Xenopus_ WISH was imaged with a Zeiss SteREO Discovery.V12 or if sections
were made with a Zeiss Axioskop 2 mot plus each equipped with an AxioCam HRc in combination with AxioVision 4.7. Fluorescence imaging of _Xenopus_ was performed on a Zeiss AxioObserver with
a LSM700 and Zeiss ZEN black software. CBF was documented with a Zeiss Axioskop 2 mot plus equipped with a high-speed Hamamatsu video camera Orca flash 4.0 and Zeiss ZEN blue. CGF analysis
was done with a Zeiss Axioskop 2 mot plus combined with a Zeiss AxioCam HSm camera and Zeiss AxioVision 4.7. CBF and CGF was assessed using the ImageJ plugin Particle Tracker56. All
trajectories were further analysed using a custom-made program written in RStudio which calculated the velocity of each particle track57. DATA AVAILABILITY The data generated or analysed
during this study are included in this article (and its Supplementary Information files). The full mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via
the PRIDE58 partner repository with the dataset identifier PXD022420. REFERENCES * Takeda, S. & Narita, K. Structure and function of vertebrate cilia, towards a new taxonomy.
_Differentiation_ 83, S4-11 (2012). Article CAS PubMed Google Scholar * Berbari, N. F., Connor, A. K. O., Haycraft, C. J. & Yoder, B. K. The primary cilium as a complex signaling
review center. _Curr. Biol._ 19, R526–R535 (2009). Article CAS PubMed PubMed Central Google Scholar * Gerdes, J. M., Davis, E. E. & Katsanis, N. The vertebrate primary cilium in
development, homeostasis, and disease. _Cell_ 137, 32–45 (2009). Article CAS PubMed PubMed Central Google Scholar * Afzelius, B. A. & Eliasson, R. Male and female infertility
problems in the immotile-cilia syndrome. _Eur. J. Respir. Dis. Suppl._ 127, 144–147 (1983). CAS PubMed Google Scholar * Stannard, W. & O’Callaghan, C. Ciliary function and the role of
cilia in clearance. _J. Aerosol. Med._ 19, 110–115 (2006). Article CAS PubMed Google Scholar * Jacquet, B. V. _et al._ FoxJ1-dependent gene expression is required for differentiation of
radial glia into ependymal cells and a subset of astrocytes in the postnatal brain. _Development_ 136, 4021–4031 (2009). Article CAS PubMed PubMed Central Google Scholar * Takeda, S.
_et al._ Left–right asymmetry and kinesin superfamily protein KIF3A: New insights in determination of laterality and mesoderm induction by kif3A−/− mice analysis. _J. Cell Biol._ 145,
825–836 (1999). Article CAS PubMed PubMed Central Google Scholar * Nonaka, S. _et al._ Randomization of left–right asymmetry due to loss of nodal cilia generating leftward flow of
extraembryonic fluid in mice lacking KIF3B motor protein. _Cell_ 95, 829–837 (1998). Article CAS PubMed Google Scholar * Nonaka, S. _et al._ De Novo formation of left–right asymmetry by
posterior tilt of nodal cilia. _PLoS Biol._ 3, e268 (2005). Article PubMed PubMed Central CAS Google Scholar * Banizs, B. _et al._ Dysfunctional cilia lead to altered ependyma and
choroid plexus function, and result in the formation of hydrocephalus. _Development_ 132, 5329–5339 (2005). Article CAS PubMed Google Scholar * Spassky, N. _et al._ Adult ependymal cells
are postmitotic and are derived from radial glial cells during embryogenesis. _Neuroscience_ 25, 10–18 (2005). Article CAS PubMed PubMed Central Google Scholar * Lee, L. Riding the
wave of ependymal cilia: Genetic susceptibility to hydrocephalus in primary ciliary dyskinesia. _J. Neurosci. Res._ 91, 1117–1132 (2013). Article CAS PubMed Google Scholar * Jain, R. _et
al._ Temporal relationship between primary and motile ciliogenesis in airway epithelial cells. _Am. J. Respir. Cell Mol. Biol._ 43, 731–739 (2010). Article CAS PubMed PubMed Central
Google Scholar * Lyons, R. A., Saridogan, E. & Djahanbakhch, O. The reproductive significance of human Fallopian tube cilia. _Hum. Reprod. Update_ 12, 363–372 (2006). Article CAS
PubMed Google Scholar * Brown, J. M. & Witman, G. B. Cilia and Diseases. _Bioscience_ 64, 1126–1137 (2014). Article PubMed PubMed Central Google Scholar * Waters, A. M. &
Beales, P. L. Ciliopathies: An expanding disease spectrum. _Pediatr. Nephrol._ 26, 1039–1056 (2011). Article PubMed PubMed Central Google Scholar * Fliegauf, M., Benzing, T. & Omran,
H. When cilia go bad: Cilia defects and ciliopathies. _Nat. Rev. Mol. Cell Biol._ 8, 880–893 (2007). Article CAS PubMed Google Scholar * Hildebrandt, F., Benzing, T. & Katsanis, N.
Ciliopathies. _N. Engl. J. Med._ 364, 1533–1543 (2011). Article CAS PubMed PubMed Central Google Scholar * Nigg, E. A. & Raff, J. W. Centrioles, centrosomes, and cilia in health and
disease. _Cell_ 139, 663–678 (2009). Article CAS PubMed Google Scholar * Praveen, K., Davis, E. E. & Katsanis, N. Unique among ciliopathies: Primary ciliary dyskinesia, a motile
cilia disorder. _F1000Prime Rep._ 7, 36 (2015). Article PubMed PubMed Central CAS Google Scholar * Lee, L. & Ostrowski, L. E. Motile cilia genetics and cell biology: Big results
from little mice. _Cell. Mol. Life Sci._ 78, 769–797 (2021). Article CAS PubMed Google Scholar * Brody, S. L., Yan, X. H., Wuerffel, M. K., Song, S. K. & Shapiro, S. D. Ciliogenesis
and left-right axis defects in forkhead factor HFH-4-null mice. _Am. J. Respir. Cell Mol. Biol._ 23, 45–51 (2000). Article CAS PubMed Google Scholar * Alten, L. _et al._ Differential
regulation of node formation, nodal ciliogenesis and cilia positioning by Noto and Foxj1. _Development_ 139, 1276–1284 (2012). Article CAS PubMed Google Scholar * Chen, J., Knowles, H.
J., Hebert, J. L. & Hackett, B. P. Mutation of the mouse hepatocyte nuclear factor/forkhead homologue 4 gene results in an absence of cilia and random left-right asymmetry. _J. Clin.
Invest._ 102, 1077–1082 (1998). Article CAS PubMed PubMed Central Google Scholar * Stubbs, J. L., Oishi, I., Izpisúa Belmonte, J. C. & Kintner, C. The forkhead protein Foxj1
specifies node-like cilia in Xenopus and zebrafish embryos. _Nat. Genet._ 40, 1454–1460 (2008). Article CAS PubMed PubMed Central Google Scholar * Yu, X., Ng, C. P., Habacher, H. &
Roy, S. Foxj1 transcription factors are master regulators of the motile ciliogenic program. _Nat. Genet._ 40, 1445–1453 (2008). Article CAS PubMed Google Scholar * Vij, S. _et al._
Evolutionarily ancient association of the FoxJ1 transcription factor with the motile ciliogenic program. _PLoS Genet._ 8, e1003019–e1003112 (2012). Article CAS PubMed PubMed Central
Google Scholar * Fritzenwanker, J. H., Gerhart, J., Freeman, R. M. & Lowe, C. J. The Fox/Forkhead transcription factor family of the hemichordate _Saccoglossus kowalevskii_. _EvoDevo_
5, 17 (2014). Article PubMed PubMed Central Google Scholar * Stauber, M. _et al._ Identification of FOXJ1 effectors during ciliogenesis in the foetal respiratory epithelium and embryonic
left-right organiser of the mouse. _Dev. Biol._ 423, 170–188 (2017). Article CAS PubMed Google Scholar * Weidemann, M. _et al._ CFAP157 is a murine downstream effector of FOXJ1 that is
specifically required for flagellum morphogenesis and sperm motility. _Development_ 143, 4736–4748 (2016). Article CAS PubMed Google Scholar * Stauber, M. _et al._ 1700012B09Rik, a FOXJ1
effector gene active in ciliated tissues of the mouse but not essential for motile ciliogenesis. _Dev. Biol._ 429, 186–199 (2017). Article CAS PubMed Google Scholar * Beckers, A. _et
al._ The evolutionary conserved FOXJ1 target gene Fam183b is essential for motile cilia in Xenopus but dispensable for ciliary function in mice. _Sci. Rep._ 8, 14678 (2018). Article ADS
PubMed PubMed Central CAS Google Scholar * Rachev, E. _et al._ CFAP43 modulates ciliary beating in mouse and Xenopus. _Dev. Biol._ 459, 109–125 (2020). Article CAS PubMed Google
Scholar * Beckers, A. _et al._ The FOXJ1 target Cfap206 is required for sperm motility, mucociliary clearance of the airways and brain development. _Development_ 147(21), dev188052 (2020).
* Pazour, G. J., Agrin, N., Leszyk, J. & Witman, G. B. Proteomic analysis of a eukaryotic cilium. _J. Cell Biol._ 170, 103–113 (2005). Article CAS PubMed PubMed Central Google
Scholar * Ma, M. _et al._ Structure of the decorated ciliary doublet microtubule. _Cell_ 179, 909-922.e12 (2019). Article CAS PubMed PubMed Central Google Scholar * Austin-Tse, C. _et
al._ Zebrafish ciliopathy screen plus human mutational analysis identifies C21orf59 and CCDC65 defects as causing primary ciliary dyskinesia. _Am. J. Hum. Genet._ 93, 672–686 (2013). Article
CAS PubMed PubMed Central Google Scholar * Choksi, S. P., Babu, D., Lau, D., Yu, X. & Roy, S. Systematic discovery of novel ciliary genes through functional genomics in the
zebrafish. _Development_ 141, 3410–3419 (2014). Article CAS PubMed PubMed Central Google Scholar * Geremek, M. _et al._ Sequence analysis of 21 genes located in the Kartagener syndrome
linkage region on chromosome 15q. _Eur. J. Hum. Genet._ 16, 688–695 (2008). Article CAS PubMed Google Scholar * Osinka, A. _et al._ Ciliary proteins: Filling the gaps. Recent advances in
deciphering the protein composition of motile ciliary complexes. _Cells_ 8(7), 730 (2019). * Sanders, A. A. _et al._ KIAA0556 is a novel ciliary basal body component mutated in Joubert
syndrome. _Genome Biol._ 16, 293 (2015). Article PubMed PubMed Central CAS Google Scholar * de Vries, W. N. _et al._ Expression of Cre recombinase in mouse oocytes: a means to study
maternal effect genes. _Genesis_ 26, 110–112 (2000). Article PubMed Google Scholar * Gene Ontology Consortium. The gene ontology resource: Enriching a GOld mine. _Nuc. Acids Res._ 49,
D325–D334 (2021). Article CAS Google Scholar * Ashburner, M. _et al._ Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. _Nat. Genet._ 25, 25–29 (2000).
Article CAS PubMed PubMed Central Google Scholar * El-Brolosy, M. A. & Stainier, D. Y. R. Genetic compensation: A phenomenon in search of mechanisms. _PLoS Genet._ 13, e1006780
(2017). Article PubMed PubMed Central CAS Google Scholar * El-Brolosy, M. A. _et al._ Genetic compensation triggered by mutant mRNA degradation. _Nature_
https://doi.org/10.1038/s41586-019-1064-z (2019). Article PubMed PubMed Central Google Scholar * Ma, Z. _et al._ PTC-bearing mRNA elicits a genetic compensation response via Upf3a and
COMPASS components. _Nature_ 568, 259–263 (2019). Article ADS CAS PubMed Google Scholar * Ichikawa, M. & Bui, K. H. Microtubule inner proteins: A meshwork of luminal proteins
stabilizing the doublet microtubule. _BioEssays_ 40, 1700209 (2018). Article CAS Google Scholar * Stoddard, D. _et al._ Tetrahymena RIB72A and RIB72B are microtubule inner proteins in the
ciliary doublet microtubules. _Mol. Biol. Cell_ 29, 2566–2577 (2018). Article CAS PubMed PubMed Central Google Scholar * Owa, M. _et al._ Inner lumen proteins stabilize doublet
microtubules in cilia and flagella. _Nat. Commun._ 10, 1143–1210 (2019). Article ADS PubMed PubMed Central CAS Google Scholar * Rodríguez, C. I. _et al._ High-efficiency deleter mice
show that FLPe is an alternative to Cre-loxP. _Nat. Genet._ 25, 139–140 (2000). Article PubMed Google Scholar * Rudat, C. _et al._ Upk3b is dispensable for development and integrity of
urothelium and mesothelium. _PLoS ONE_ 9, e112112 (2014). Article ADS PubMed PubMed Central CAS Google Scholar * Moorman, A. F., Houweling, A. C., de Boer, P. A. & Christoffels, V.
M. Sensitive nonradioactive detection of mRNA in tissue sections: Novel application of the whole-mount in situ hybridization protocol. _J. Histochem. Cytochem._ 49, 1–8 (2001). Article CAS
PubMed Google Scholar * Kawai, J. & Hayashizaki, Y. DNA book. _Genome Res._ 13, 1488–1495 (2003). Article CAS PubMed PubMed Central Google Scholar * You, Y. & Brody, S. L.
Culture and differentiation of mouse tracheal epithelial cells. _Methods Mol. Biol._ 945, 123–143 (2013). Article PubMed CAS Google Scholar * Sbalzarini, I. F. & Koumoutsakos, P.
Feature point tracking and trajectory analysis for video imaging in cell biology. _J. Struct. Biol._ 151, 182–195 (2005). Article CAS PubMed Google Scholar * Vick, P. _et al._ Flow on
the right side of the gastrocoel roof plate is dispensable for symmetry breakage in the frog _Xenopus laevis_. _Dev. Biol._ 331, 281–291 (2009). Article CAS PubMed Google Scholar *
Vizcaíno, J. A. _et al._ 2016 update of the PRIDE database and its related tools. _Nuc. Acids Res._ 44, D447–D456 (2016). Article CAS Google Scholar Download references ACKNOWLEDGEMENTS
We thank S. Brody (Washington University, St. Louis) for _Foxj1_-mutant mice. This work was financially supported by the DFG (GO 449/14-1 and BL285/14-1) and by the DFG Cluster of Excellence
“REBIRTH” to AG. TO was supported by a Ph.D. fellowship from the Landesgraduiertenförderung Baden-Württemberg. MU was supported by the Tistou & Charlotte Kerstan Stiftung. PW is funded
by the Deutsche Forschungsgemeinschaft (DFG) under the Emmy Noether Programme (Grant WA3365/2-1) and under Germany’s Excellence Strategy (CIBSS-EXC-2189-Project ID 390939984). RNA-Sequencing
data used or referred to in this publication were generated by the Research Core Unit Genomics (RCUG) at Hannover Medical School. For access to the CASA instruments and their help we thank
D. Conrad, C. Schippert and P. Hillemanns (Department of Gynaecology and Obstetrics at Hannover Medical School). FUNDING Open Access funding enabled and organized by Projekt DEAL. AUTHOR
INFORMATION Author notes * Magdalena Maria Brislinger Present address: Renal Division, Department of Medicine, University Hospital Freiburg, Freiburg University Faculty of Medicine &
CIBSS-Centre for Integrative Biological Signalling Studies, University of Freiburg, Habsburger Str. 49, 79104, Freiburg, Germany * Leonie Alten Present address: Twist Bioscience, 681 Gateway
Blvd South, South San Francisco, CA, 94080, USA * Elisabeth Kremmer Present address: Department of Biology II, Ludwig-Maximilians University, Großhaderner Straße 2, 82152, Martinsried,
Germany * Adina Przykopanski Present address: Institute for Toxicology, OE 5340, Hannover Medical School, Carl-Neuberg-Str. 1, 30625, Hannover, Germany AUTHORS AND AFFILIATIONS * Institute
for Molecular Biology, OE5250, Hannover Medical School, Carl-Neuberg-Str. 1, 30625, Hannover, Germany Anja Beckers, Karin Schuster-Gossler, Leonie Alten, Adina Przykopanski, Katrin Serth
& Achim Gossler * Institute of Biology, University of Hohenheim, Garbenstraße 30, 70593, Stuttgart, Germany Franziska Fuhl, Tim Ott, Magdalena Maria Brislinger & Martin Blum *
Institute of Ophthalmic Research, Center for Ophthalmology, University of Tübingen, Elfriede-Aulhorn-Strasse 7, 72076, Tübingen, Germany Karsten Boldt & Marius Ueffing * Renal Division,
Department of Medicine, University Hospital Freiburg, Freiburg University Faculty of Medicine & CIBSS-Centre for Integrative Biological Signaling Studies, University of Freiburg,
Habsburger Str. 49, 79104, Freiburg, Germany Peter Walentek * Institute of Functional and Applied Anatomy, Research Core Unit Electron Microscopy, OE8840, Hannover Medical School,
Carl-Neuberg-Str. 1, 30625, Hannover, Germany Jan Hegermann * Institute of Molecular Immunology, Helmholtz Zentrum München, German Research Center for Environmental Health (GmbH), Core
Facility Monoclonal Antibodies, Marchioninistr. 25, 81377, München, Germany Elisabeth Kremmer Authors * Anja Beckers View author publications You can also search for this author inPubMed
Google Scholar * Franziska Fuhl View author publications You can also search for this author inPubMed Google Scholar * Tim Ott View author publications You can also search for this author
inPubMed Google Scholar * Karsten Boldt View author publications You can also search for this author inPubMed Google Scholar * Magdalena Maria Brislinger View author publications You can
also search for this author inPubMed Google Scholar * Peter Walentek View author publications You can also search for this author inPubMed Google Scholar * Karin Schuster-Gossler View author
publications You can also search for this author inPubMed Google Scholar * Jan Hegermann View author publications You can also search for this author inPubMed Google Scholar * Leonie Alten
View author publications You can also search for this author inPubMed Google Scholar * Elisabeth Kremmer View author publications You can also search for this author inPubMed Google Scholar
* Adina Przykopanski View author publications You can also search for this author inPubMed Google Scholar * Katrin Serth View author publications You can also search for this author inPubMed
Google Scholar * Marius Ueffing View author publications You can also search for this author inPubMed Google Scholar * Martin Blum View author publications You can also search for this
author inPubMed Google Scholar * Achim Gossler View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS Conceptualisation: M.B., A.B., A.G., T.O.
Methodology: A.B., K.S.G., J.H., K.B., F.F., M.M.B., T.O. Reagents: E.K. Formal analysis: A.B., K.S.G., T.O., F.F., J.H., K.B., K.S., P.W., M.B., A.G. Investigation: A.B., K.S.G., M.M.B.,
A.P., T.O., F.F., J.H., K.B., K.S., L.A. Writing—original draft: M.B., A.G., A.B., P.W. Writing—review and final editing: all authors. Supervision: M.B., A.G., M.U. Project administration:
M.B., A.G. Funding acquisition: M.B., A.G., M.U., P.W. CORRESPONDING AUTHORS Correspondence to Martin Blum or Achim Gossler. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no
competing interests. ADDITIONAL INFORMATION PUBLISHER'S NOTE Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
SUPPLEMENTARY INFORMATION SUPPLEMENTARY INFORMATION 1. SUPPLEMENTARY INFORMATION 2. SUPPLEMENTARY INFORMATION 3. RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative
Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the
original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in
the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your
intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence,
visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Beckers, A., Fuhl, F., Ott, T. _et al._ The highly conserved FOXJ1 target
CFAP161 is dispensable for motile ciliary function in mouse and _Xenopus_. _Sci Rep_ 11, 13333 (2021). https://doi.org/10.1038/s41598-021-92495-3 Download citation * Received: 27 November
2020 * Accepted: 08 June 2021 * Published: 25 June 2021 * DOI: https://doi.org/10.1038/s41598-021-92495-3 SHARE THIS ARTICLE Anyone you share the following link with will be able to read
this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative