- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT Heat shock factor 1 (HSF1) initiates a broad transcriptional response to proteotoxic stress while also mediating a cancer-specific transcriptional program. HSF1 is thought to be
regulated by molecular chaperones, including Heat Shock Protein 90 (HSP90). HSP90 is proposed to sequester HSF1 in unstressed cells, but visualization of this interaction _in vivo_ requires
protein crosslinking. In this report, we show that HSP90 binding to HSF1 depends on HSP90 conformation and is only readily visualized for the ATP-dependent, N-domain dimerized chaperone, a
conformation only rarely sampled by mammalian HSP90. We have used this mutationally fixed conformation to map HSP90 binding sites on HSF1. Further, we show that ATP-competitive, N-domain
targeted HSP90 inhibitors disrupt this interaction, resulting in the increased duration of HSF1 occupancy of the _hsp70_ promoter and significant prolongation of both the constitutive and
heat-induced HSF1 transcriptional activity. While our data do not support a role for HSP90 in sequestering HSF1 monomers to suppress HSF1 transcriptional activity, our findings do identify a
noncanonical role for HSP90 in providing dynamic modulation of HSF1 activity by participating in removal of HSF1 trimers from heat shock elements in DNA, thus terminating the heat shock
response. SIMILAR CONTENT BEING VIEWED BY OTHERS REGULATORY INTER-DOMAIN INTERACTIONS INFLUENCE HSP70 RECRUITMENT TO THE DNAJB8 CHAPERONE Article Open access 11 February 2021 REVERSIBLE
PHASE SEPARATION OF HSF1 IS REQUIRED FOR AN ACUTE TRANSCRIPTIONAL RESPONSE DURING HEAT SHOCK Article 07 March 2022 TRANSCRIPTIONAL RESPONSES OF CANCER CELLS TO HEAT SHOCK-INDUCING STIMULI
INVOLVE AMPLIFICATION OF ROBUST HSF1 BINDING Article Open access 16 November 2023 INTRODUCTION Heat shock factor 1 (HSF1) is an evolutionarily conserved transcription factor that initiates
the cytoprotective heat shock response (HSR). Found throughout the eukaryotic kingdom, HSF1 allows for the cellular adaptation to proteotoxic stress1. Through an incompletely defined
mechanism, mammalian HSF1 monomers in cytosol are activated and form trimers, translocate into the nucleus, and bind sequences of DNA known as heat shock elements (HSE), ideally represented
as nGAAnnTTCnnGAAn2,3. Throughout this process HSF1 is heavily post-translationally modified and interacts with numerous cellular components. The binding of HSF1 trimers to HSE induces the
transcription of a specialized set of genes known as molecular chaperones while also repressing the expression of other genes4,5, although the repressive effect of HSF1 is controversial6.
Once expressed, these molecular chaperones (or Heat Shock Proteins, HSPs) act to stabilize the three-dimensional structure of numerous cellular proteins, thus helping to maintain cellular
proteostasis. HSP90 and HSP70 are ATP-dependent HSPs that interact with a large sector of the eukaryotic proteome while also modulating HSF1 transcriptional activity7,8,9. The relationship
between HSF1 and one or more components of the cellular proteostasis network is thought to represent a primary axis in the control of the HSR7,10. In cancer, HSF1 enables malignant cell
growth, is overexpressed in a number of tumor types, and is associated with poor prognosis11,12,13. Although HSF1 does not initiate oncogenic transformation, tumors become addicted to HSF1
activity as their microenvironments become increasingly toxic and as they require higher levels of HSPs to maintain proteostasis14. Moreover, many oncogenes that drive tumorigenesis are
metastable and rely on HSPs to sustain their activity. This is particularly true for mutated or overexpressed kinases and transcription factors that interact with HSP9015. HSF1 also promotes
a cancer-specific transcriptional program that supports malignancy through the expression of genes for proliferation, anabolic metabolism, metastasis and apoptosis prevention. Comprised of
over 500 genes, this cancer-specific HSF1 transcriptome is associated with poor clinical outcomes11. The human _HSF1_ gene is encoded on chromosome 8q24 by 14 exons that produce two splice
variants. The largest variant, which is described in this report, is translated into 529 amino acids. HSF1 has a predicted molecular weight of 57 kDa, yet migrates at approximately 75 kDa on
SDS-PAGE due to a large number of post-translational modifications (PTMs), including phosphorylation, acetylation and sumoylation16,17. The overall structure of HSF1 is mostly disordered
except for the evolutionarily conserved N-terminal DNA-binding domain (DBD) that forms a winged helix-turn-helix structure18,19. The rest of HSF1 is predicted not to maintain a stable
tertiary structure, a feature observed for many proteins involved in transcription and cellular regulation20. Following the DBD and a linker region is the set of heptad repeats (HR-A/B) that
form the leucine zippers that allow for HSF1 trimerization21. Adjacent to the HR-A/B, the unstructured regulatory domain (RD) is the molecular region understood to be capable of sensing
heat and initiating the HSR22. The RD contains numerous phosphorylation sites23 and functions, along with a portion of HR-A/B, to repress the transcriptional activity of NF-IL624. Another
heptad repeat (HR-C), C-terminal to the RD, is understood to sequester HR-A/B in an intramolecular interaction that suppresses spontaneous HSF1 trimerization21. Most recently, Hentze _et
al_. have shown that HSP90β interacts with the HR-C domain in solution to promote its temperature-sensitive dissociation from HR-A/B, thus reducing the temperature at which HSF1 is able to
trimerize25. The C-terminal transactivation domain (TAD) is highly disordered and is required for robust HSF1 transcriptional activity in response to stress26. The TAD is also a site of
HSP70 binding, which negatively regulates HSF1 transcriptional activity27. While the interaction of HSP70 with HSF1 in unstressed cells is readily observed by co-immunoprecipitation27,
interaction of endogenous HSP90 and HSF1 is not readily detectable and requires cross-linking to stabilize the association7. These data argue against a frequently cited model postulating
sequestration of HSF1 monomers by HSP90 under non-stress conditions (7), but they would support such a role for HSP70, as has recently been proposed in yeast28. Chaperone binding to HSF1
monomers is thought to prevent spontaneous trimerization. In such a model, HSF1 is released from chaperone suppression and allowed to trimerize when cells experience severe proteotoxic
stress that generates a large population of denatured client proteins and an increased demand for chaperones29. Other studies have shown that HSP90 can also bind the trimerized DNA-bound
state of HSF1 and that this interaction attenuates HSF1 transcriptional activity8. Thus, HSP90 has variously been proposed to function as a sensor, repressor and/or attenuator of the HSR.
Determining the mechanism by which HSP90 affects the HSR is of medical significance. Currently, HSP90 is the only HSP whose targeting by small molecules has been extensively evaluated in the
clinic as a strategy to treat cancer30. However, although HSP90 inhibitors have shown great promise in pre-clinical studies due to their ability to disrupt proteostasis and to promote the
degradation of oncogenic kinases and other pro-growth components31, their single agent activity in patients has not been impressive32,33. This discrepancy has been ascribed, in part, to the
observation that ATP-competitive HSP90 inhibitors promote the HSR and result in the increased expression of HSP70, HSP27, and other pro-survival components34,35, possibly offsetting any
anti-cancer benefit of these drugs. Moreover, HSP90 inhibitors may enhance the recently identified cancer-specific HSF1 transcriptional program11. How this is mechanistically achieved in the
absence of robust HSP90/HSF1 interaction has remained obscure. In this report, we define the interactions of HSF1 with both nuclear-cytosolic paralogs of HSP90, stress-inducible HSP90α and
constitutively expressed HSP90β, and with endogenous HSP70. Remarkably, we readily observed interaction of HSF1 with mutant HSP90 trapped in an ATP-dependent “closed” conformation (unable to
hydrolyze ATP), and we found this interaction to be disrupted by N-terminal HSP90 inhibitors that are currently being clinically evaluated. To determine the domains in HSF1 that bind HSP90
and HSP70 we made a series of HSF1 deletion mutants and analyzed their interactions by immunoprecipitation-western blot. We observed that the binding of HSP90α, HSP90β and HSP70 uniquely map
across HSF1. This was complemented by analysis of the effects of these deletion constructs on HSF1 trimerization and HSR reporter activity. Finally, we found that HSP90 inhibitors disrupt
association of “closed” HSP90 proteins with HSF1, while concomitantly increasing the duration of HSF1 binding to _hsp70_ promoter and extending the duration of heat-induced HSF1
transcriptional activity. While our data do not support a role for HSP90 in sequestering HSF1 monomers, our findings reveal that HSP90 inhibitors interfere with a noncanonical role for HSP90
in providing dynamic modulation of HSF1 activity by removing HSF1 trimers from heat shock elements in DNA. RESULTS WILD TYPE HSF1 READILY INTERACTS WITH N-DOMAIN DIMERIZED (“CLOSED”
CONFORMATION) HSP90 Previous studies have suggested that the intracellular interaction of HSF1 and HSP90 is weak and transient, and requires chemical crosslinking for visualization7. We
showed recently that the strength of this interaction may be influenced by HSP90 conformation36. Binding of ATP to N-terminal domains of HSP90 promotes their transient dimerization and
induces a “closed” conformation that is poised for ATP hydrolysis. ATP binding and hydrolysis, in turn, drive the structural rearrangements in HSP90 that support chaperone function. Upon
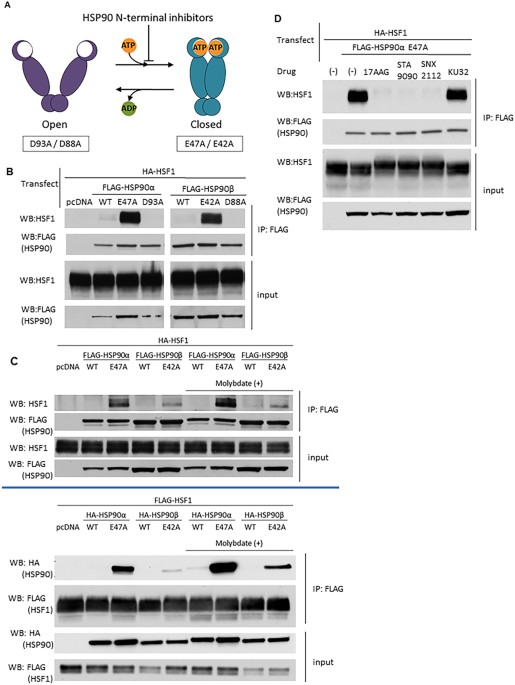
release of ADP, or in the presence of ATP-competitive HSP90 inhibitors, HSP90 N-domains revert to an undimerized state (“open” conformation) (Fig. 1A). Mutation of key residues within the
N-terminal domain of HSP90 stabilize either the “closed” (HSP90α E47A, HSP90β E42A; allow ATP binding but prevent hydrolysis) or “open” (HSP90α D93A, HSP90β D88A; prevent ATP binding)
conformations. We co-transfected HEK293 cells with plasmids encoding FLAG-tagged wild type or conformationally restricted HSP90 proteins together with wild type HA-HSF1, harvested the cells
the following day, and observed the protein-protein interactions by anti-FLAG immunoprecipitation-western blot (IP-WB) analysis (Fig. 1B,C). The relative expression of HSF1 in transfected
and untransfected cells is shown in Supplemental Fig. 1A. We found that wild type HSF1 strongly interacts, without the need for crosslinking, with both “closed” HSP90 mutants but not with
the pair of “open” HSP90 mutants. Importantly, the results with transfected HSF1 were confirmed by co-immunoprecipitation of “closed” HSP90 with endogenous HSF1 in HEK293 cells (Supplemental
Fig. 1B). As previously reported7, interaction of wild type HSP90, which samples all available conformational states, with full-length HSF1 is not detectable in the absence of protein
crosslinking. This result is consistent with the observation that human HSP90, in contrast to the yeast chaperone, is most likely to be found in the “open” conformation, even when ATP is
bound37. Moreover, we observed that HSP90α E47A bound more avidly to HSF1 than did HSP90β E42A, suggesting that the stress-inducible HSP90α isoform may be a stronger negative regulator of
HSF1 activity compared to the constitutively expressed HSP90β isoform, as previously proposed36. To further assess the interaction between wild type HSF1 and the “closed” HSP90 mutants, we
repeated the reciprocal experiment in the absence or presence of the anion molybdate (MoO4−2) Molybdate has been reported to stabilize a high affinity client binding state of HSP9038. Target
proteins were pulled down via either FLAG-HSP90 or FLAG-HSF1 (Fig. 1C). Although, the HSF1/ “closed” HSP90 interaction was confirmed, molybdate only moderately increased the interaction of
the “closed” HSP90 mutants with no discernable effect on interaction of wild type HSP90 with HSF1, indicating that HSP90 interacts with HSF1 in a manner that is likely unique when compared
to other clients such as protein kinases38,39. Administration of HSP90 inhibitors often results in apparent induction or augmentation of the HSR. Currently, there are two general classes of
inhibitors. The more clinically advanced N-terminal inhibitors competitively bind to the ATP pocket and prevent or disrupt nucleotide-dependent N-domain dimerization40. These inhibitors
include 17AAG (tanespimycin), STA9090 (ganetespib) and SNX2112, among others30. A second class of inhibitors, based on the antibiotic novobiocin, binds a putative nucleotide-binding domain
in the C-terminal portion of HSP90 (adjacent to the C-terminal dimerization domain) and inhibits the chaperone in a distinct manner41. The synthetic novobiocin derivative KU32 is
representative of this class of inhibitors42. To explore how N- and C-terminal inhibitors might affect HSF1/HSP90 interaction, we co-transfected HEK293 cells with wild type HA-HSF1 and
FLAG-HSP90α E47A, briefly treated cells with inhibitors the following day, harvested and analyzed protein-protein interactions by IP-WB (Fig. 1D, Supplemental Fig. 1C). Each of the three
N-terminal inhibitors disrupted the interaction between HSF1 and HSP90α E47A, consistent with their disruption of N-domain dimerization. In contrast, KU32 did not disrupt the interaction
between HSF1 and “closed” HSP90, indicating that this is a unique property of ATP-competitive, N-terminal HSP90 inhibitors. MAPPING OF HSP90 AND HSP70 INTERACTIONS ACROSS HSF1 To investigate
the interactions of HSP90 and HSP70 with HSF1 in the absence of a crosslinking agent, we constructed and transiently expressed in HEK293 cells a series of HSF1 truncation and internal
deletion mutants, and we analyzed them for chaperone interaction by IP-WB (Fig. 2A,B). Of note, N195, an N-terminal truncation mutant lacking the DBD and most of the HR-A/B domain, expressed
very poorly (see blot in Fig. 2B) and data obtained using this mutant should be considered in this context. These HSF1 constructs were used to initially query the interaction of HSP90 and
HSP70 across the TAD, HR-C, RD, HR-A/B and DBD domains of HSF1 (Supplemental Fig. 2A,B). Based on these preliminary data obtained with the closed HSP90 mutants, a subpanel of HSF1 truncation
and deletion mutants were selected to assess association of HSP90α E47A. Pulldowns were repeated in triplicate, resulting blots were subjected to densitometry to generate mean values +/−
standard deviations and p values to assess significance (compared to association of HSP90α E47A with full length HSF1) were obtained. These data are shown in Fig. 2, panels C and D. Analysis
of HSF1 C-terminal truncations revealed a distinct interaction motif for “closed” HSP90 E47A (Fig. 2C). The “closed” mutant bound most intensely to HSF1 proteins containing the HR-A/B
domain and a portion of the heat-sensing RD, but lacking the HR-C and TAD. The clear difference in HSP90 interaction between the C442 and C383 HSF1 truncation mutants suggests that presence
of the HSF1 HR-C + TAD domain inhibits HSP90 interaction. Although HSP90 E47A bound weakly to more extensive C-terminal truncations, binding was not significantly different from wild type
HSF1. The dual importance of both the HR-A/B and RD domains for HSP90 binding is supported by the observation that neither the isolated HR-A/B domain (N120-C205, Fig. 2D and Supplemental
Fig. 2B) nor the isolated RD domain (N262-C383, Supplemental Fig. 2B) display binding to HSP90. Importantly, deletion of the HR-C domain dramatically improves the binding of both “closed”
and endogenous HSP90, suggesting that HR-C intermolecular binding to HR-A/B (characteristic of inactive HSF1 monomers) interferes with HSP90 binding and providing further evidence that is
inconsistent with a role for HSP90 in sequestering monomeric HSF1. Finally, the HSF1 HR-A/B internal deletion construct DEL142–169, where amino acids 142–169 were removed, bound “closed”
HSP90 while the HR-A/B internal deletion construct DEL183–214 did not (Fig. 2D and Supplemental Fig. 2D), suggesting that the C-terminal half of the HR-A/B is necessary for HSP90 binding.
While the binding pattern of endogenous HSP90 generally mirrors that of “closed” HSP90 (although, in contrast to “closed” HSP90, detectable binding of endogenous HSP90 requires deletion of
the HR-C domain, see Supplemental Fig. 2A,B), endogenous HSP90 also uniquely binds the HSF1 constructs N195 and N262 but not the equivalent HSF1 mutants lacking the HR-C and TAD domains
(N195-C383 or N262-C383, see Supplemental Fig. 2B). These findings are consistent with the interaction of endogenous HSP90 (primarily existing in an N-domain undimerized conformation) with
the HR-C domain, as recently reported by Mayer and colleagues for HSP90β. Importantly, these authors demonstrated that HSP90 binding to the HSF1 HR-C domain promotes its
temperature-sensitive dissociation from HR-A/B, thus facilitating HSF1 trimerization25. Like our data, these findings are also not consistent with a role for HSP90 in suppressing HSF1
trimerization. In contrast to the binding profile of HSP90, endogenous HSP70 interacts robustly with full-length HSF1 as well as with most of the C-terminal truncation and deletion mutants
(Supplemental Fig. 2A and B), with the exception of those mutants containing increasingly truncated RD. Endogenous HSP70 also bound poorly to N120-C205 (isolated HR-A/B domain) despite ample
expression of this HSF1 protein fragment. Taken together, these data identify the RD as being sufficient for HSP70 binding in unstressed cells, although HSP70 binding to other domains of
HSF1 has also been reported26,27. To further compare the role of the HR-A/B domain in HSF1 trimerization and in HSP90 interaction, we made a set of alanine scanning mutants (145A4, 151A5,
161A5) between amino acids142–169 (see Methods) and compared them in the experiments below with the internal deletion constructs DEL142–169 and DEL183–214. Using IP-WB analysis of
co-transfected WT HA-HSF1 and mutant FLAG-HSF1, we observed that the HR-A/B domain alanine mutants did not disrupt HSF1 intermolecular interaction, in distinct contrast to the HR-A/B
deletion mutants DEL142–169 and DEL182–214 (Supplemental Fig. 2C). In contrast, these alanine mutants bound “closed” HSP90α E47A to a greater extent compared to WT HSF1 (but similarly to
DEL142–169 (Supplemental Fig. 2D and Fig. 2D). These data are consistent with the hypothesis that the HR-A/B region contributing to HSF1 trimerization is distinct from the HR-A/B region that
is required for HSP90 binding. Further, our findings emphasize that the presence or absence of HSP90 binding has no impact on HSF1 trimerization (see Fig. 3B,D and Supplemental Fig. 5).
Finally, the enhanced HSP90 binding of the HSF1 truncation and deletion mutants shown in Fig. 2C,D does not involve possible heterotrimerization of the transfected HSF1 constructs with
endogenous HSF1, as similar results are obtained when the HSF1 truncation mutants are transfected into HEK293 cells stably knocked down for endogenous HSF1 (Supplemental Fig. 2E). THE TAD
AND DBD ARE ESSENTIAL FOR HSF1 TRANSCRIPTIONAL ACTIVITY WHILE THE HR-A/B DOMAIN IS REQUIRED FOR DOMINANT NEGATIVE ACTIVITY OF TRUNCATED HSF1 To further explore potential relationships
between the ability of HSF1 to (1) form intermolecular trimers, (2) activate transcription, and (3) interact with HSP90, we examined the impact of HSF1 N/C-truncations and internal deletions
on _hsp70_ promoter-reporter activity as a readout of the HSR, and we complemented these observations with IP-WB analysis of HSF1 trimerization and HSF1-HSP90 interaction. Overexpression of
WT HSF1 increased _hsp70_-reporter activity in response to heat shock by nearly 2-fold above a pcDNA control. Brief pretreatment of cells with 17AAG prior to heat shock increased
_hsp70_-reporter activity in pcDNA control by 3.5-fold compared to heat shock alone, and a similar fold increase in reporter activity was observed upon 17AAG exposure prior to heat shock in
cells transfected with WT HSF1 (Fig. 3A). For reference, reporter activity in response to 17AAG alone, compared to heat shock and heat shock + 17AAG is shown in Supplemental Fig. 3A. Both
heat-induced and 17AAG + heat-induced _hsp70_-reporter activity were progressively impacted by increasing truncation of the TAD (C-terminal deletion constructs C509, C477 and C442),
consistent with the importance of this domain in modulating HSF1 activation43. Complete removal of TAD, HR-C and RD domains (constructs C383 through C205) fully repressed _hsp70_-reporter
activity even with 17AAG pretreatment, suggesting that the presence of the HR-A/B domain of these mutants is essential for dominant-negative activity, as a consequence of disrupting
endogenous HSF1 trimers. Indeed, truncation through the C-terminal portion of the HR-A/B domain (C182) restored _hsp70_-reporter response to that seen after treating pcDNA control cells with
heat and heat + 17AAG. Consistent with this result, HSF1 C182 is unable to form an intermolecular interaction with wild type HSF1, as are additional HR-A/B truncations (Fig. 3D).
Unexpectedly, further truncation (C167, C146 and C129) through the HR-A/B, increased _hsp70_-reporter activity above that of the pcDNA control, suggesting the presence of a repressing motif
located between amino acids 167 and 182 of HSF1 (Fig. 3A). Our previous observation of HSP90 binding to a series of RD-containing C-terminal truncation mutants (C383, C339, C311, Fig. 2C)
suggested that HSP90 binding might be involved in repressing HSF1-driven _hsp70_-reporter activity. To address this possibility, we used IP-WB to investigate the effect of 17AAG on the
interaction of HSP90α E47A with these HSF1 RD truncation mutants since these mutants repressed _hsp70_-reporter activity even in the presence of 17AAG (Fig. 3A). Consistent with the data in
Fig. 2C, we observed increased binding of HSP90α E47A to the RD truncation mutants C383, C339 and C311 when compared to wild type HSF1. However, upon addition of 17AAG, HSF1-HSP90α E47A
interaction was fully disrupted (Fig. 3B), while _hsp70_-reporter activity remained repressed (Fig. 3A). These data indicate that robust HSP90 interaction with HSF1 is not responsible for
the dominant negative activity of these mutants. Consistent with this hypothesis, the RD truncation mutant C287 bound poorly to HSP90α E47A, yet displayed dominant negative activity (see
Fig. 3A,B). Taken together, our findings support the hypothesis that the dominant negative activity of HSF1 C-terminal truncations retaining an intact HR-A/B domain is due to disruption of
endogenous full length HSF1 trimers necessary for transcriptional activity. Next, we assessed the effects of another set of HSF1 N/C-terminal deletion constructs on _hsp70_-reporter activity
(Fig. 3C). Here we observed that the deletion constructs N120, N120-C383, N120-C325 and N120-C287, all lacking the DBD but including an intact HR-A/B domain, repressed _hsp70_-reporter
activity below the level seen with empty plasmid, consistent with dominant negative inhibition of endogenous HSF1. Removal of the HR-A/B domain (N195, N262) relieved this repression to some
extent, even when the TAD and RD were removed (N195-C383, N195-C325, N262-C383, N262-C325). Finally, the internal HR-A/B deletion mutants DEL142–169 and DEL182–214 supported _hsp70_-reporter
activity at a level similar to that of empty plasmid, confirming that an intact HR-A/B domain is required for dominant negative activity. The statistical significance of the changes in
reporter activity can be found in Supplemental Fig. 3B. Of note, these data are in contrast to the impact of these two HR-A/B internal deletions on HSP90 binding (Fig. 2D, right two lanes).
To profile the ability of these deletion constructs to affect HSF1 intermolecular interaction (indicative of trimerization capability), we analyzed by IP-WB the interaction of co-transfected
wild type HA-HSF1 with various FLAG-HSF1 deletion mutants. We found that FLAG-tagged deletion mutants (C383, C287, C233, C205 and N120) that dominant negatively repressed _hsp70_-reporter
activity interacted efficiently (equivalent or enhanced interaction compared to FLAG-tagged wild type HSF1) with wild type HA-HSF1. In contrast, deletion mutants (C182, C167, C146, C129,
DEL142–169, DEL183–214) that did not strongly repress _hsp70_-reporter activity did not interact with WT HA-HSF1 (Fig. 3D). We excluded N195 from this analysis due to its poor expression as
shown in Figs 2B and 3D. Likewise, the dominant negative HSF1 mutants N120, N120-C383, N120-C325 and N120-C287 strongly interacted with full length HSF1, while N195-C383 (which showed no
dominant negative activity) did not (Supplemental Fig. 3C and D). Combined with our previous observations, these data further confirm that HSF1 mutant-mediated dominant negative repression
of endogenous HSF1-driven transcription depends on the ability to form an intermolecular interaction with wild type HSF1. A cartoon summarizing these data is provided in Fig. 3E. IMPACT OF
RD PHOSPHORYLATION STATE ON HSP90 INHIBITOR EFFECT ON HSF1 ACTIVITY AND MOBILITY SHIFT IN SDS-PAGE, AND ON HSF1 INTERACTION WITH HSP90 Stress-inducible phosphorylation of the HSF1 RD
coincides with acquisition of transactivation capacity, although a functional requirement of RD phosphorylation for HSF1 activation has not been demonstrated. Indeed, a recent report
dissociates RD phosphorylation from HSF1 activation and suggests that RD phosphorylation may serve as a rheostat to modulate the degree of activation23. Since N-terminally targeted HSP90
inhibitors also cause a distinct, phosphorylation-dependent mobility shift of HSF1 coincident with disrupting its association with conformationally restricted HSP90 proteins (see Fig. 1D,
Supplemental Figs 1C and 4), we wished to determine whether these drugs promoted phosphorylation of sites only within the RD and whether RD phosphorylation was a prerequisite for HSP90
inhibitor-enhanced HSF1 activity. For this purpose, we used the delta PRD (dPRD) HSF1 construct previously described, in which all phosphorylation sites within the RD have been mutated to
alanine (see Fig. 4A)23. Using the _hsp70_-reporter assay, we confirmed that dPRD HSF1 was more sensitive to heat shock, and to the combination of heat shock and 17AAG, compared to wild type
HSF1 (Fig. 4B). Unexpectedly, the mobility of HSF1 dPRD was also shifted when challenged with three N-domain HSP90 inhibitors (Fig. 4C), strongly suggesting that HSP90 inhibitor-induced
HSF1 phosphorylation is not restricted to site(s) within the RD (Supplemental Fig. 4). Importantly, lack of phosphorylation within the RD domain does not affect trimerization of either full
length HSF1 or the various HSF1 deletion mutants discussed above (Supplemental Fig. 3E). HSP90 INHIBITION INCREASES DURATION OF HSF1 BINDING TO _HSP70_ PROMOTER AND PROLONGS DURATION OF HSF1
TRANSCRIPTIONAL ACTIVITY IN RESPONSE TO HEAT SHOCK To assess the outcome of combining HSP90 inhibition with heat shock compared to either inhibitor or heat shock alone, we monitored the
extent and duration of HSF1 occupancy of the _hsp70_ promoter by chromatin immunoprecipitation (ChIP) with antibody to HSF1. HEK293 cells were pretreated for 1 hour at 37 °C with 10 μM 17AAG
or left untreated, and then exposed to heat shock (42 °C for 30 minutes). After recovery for varying times at 37 °C (no recovery, 2 hours, 4 hours, 6 hours), HSF1 ChIP was performed and
pulldown of associated _hsp70_ promoter sequences was assessed by PCR. An additional group exposed to 17AAG but in the absence of heat shock was also subjected to ChIP analysis at the
indicated times (based on recovery time after heat shock). The top panel in Fig. 5A shows a representative experiment in which the expected PCR fragments were run on an agarose gel. In the
bottom panel, PCR data reflecting the extent and duration of HSF1 occupancy of the _hsp70_ promoter were quantified by densitometry (3 independent experiments +/− standard deviation),
normalized to input and graphed as shown (Fig. 5A, values obtained in the absence of both heat shock and 17AAG pretreatment were arbitrarily set to 1). In the absence of 17AAG, HSF1 binding
to the _hsp70_ promoter was highest immediately after heat shock (no recovery). Promoter occupancy declined below baseline by 2 hours and remained below baseline for the duration of the
experiment. In contrast, 1-hour pre-exposure of cells to 17AAG prior to heat shock resulted in an increased duration of HSF1 binding to the _hsp70_ promoter (significantly above baseline at
2 hours of recovery and significantly above the heat shock only value at later time points). Importantly, the 17AAG alone samples displayed the same maximal level of HSF1 binding to _hsp70_
promoter as did both the heat shock alone samples and the heat shock + 17AAG samples. However, in contrast to cells treated with heat shock alone, HSF1 promoter occupancy did not decline as
rapidly in cells treated with 17AAG alone or in combination with heat shock. Thus, the level of HSF1 binding to DNA in these groups remained significantly above heat shock only values for
the full 6-hour recovery period (at least 4 hours longer than after exposure to heat shock alone). Additionally, we assessed HSF1 transcriptional activity in HEK293 cells after heat shock
alone, 17AAG alone, or after pre-exposure to 17AAG followed by heat shock, using the same experimental conditions as for the ChIP analysis above (Fig. 5B). For this purpose, we examined
endogenous _HSP70_ gene expression by RT-qPCR. Similar to the impact of heat shock on HSF1 promoter occupancy, relative _HSP70_ mRNA expression peaked after 2 hours of recovery from heat
shock and declined thereafter to below baseline by 6 hours. In contrast, 17AAG alone promoted a modest increase in _HSP70_ mRNA (although below the peak seen after heat shock) that remained
elevated at 2 and 6 hours. After combined 17AAG treatment and heat shock, _HSP70_ mRNA level increased on par with exposure to heat shock alone (similar values at 2 hours), but continued to
increase to a peak value at 4 hours (25% higher than the heat shock alone peak value) that was maintained at 6 hours. Importantly, the 4- and 6-hour increase in _HSP70_ mRNA after
combination treatment was significantly higher (2–2.5-fold) compared to HSP90 inhibitor alone. DISCUSSION In this study, we assessed the importance of HSP90 conformation for binding to HSF1
and we found that only the ATP-dependent “closed” (e.g., N-domain dimerized) conformational mutants E47A (HSP90α) and E42A (HSP90β) readily co-precipitate full-length HSF1, with HSP90α
displaying the stronger interaction. As HSP90α is stress-inducible and transcriptionally regulated by HSF1, this provides an appealing feedback control mechanism to attenuate the HSR,
consistent with a model first proposed nearly twenty years ago8,44. Indeed, our previous finding that transient over-expression of HSP90α reduced HSF1 transcriptional activity supports this
possibility36. In the absence of these conformationally restrictive mutations, mammalian HSP90 only rarely and transiently occupies the “closed” conformation, even in the presence of ATP,
with the bulk of the protein at steady-state occupying an N-domain undimerized conformation37. This is inconsistent with a role for HSP90 in binding to and sequestering HSF1 monomers to
prevent trimerization, as has also been proposed7. To identify chaperone binding sites in HSF1, we used a series of Flag-tagged HSF1 truncation and internal deletion mutants to interrogate
binding of HSP90 and HSP70, including the binding of transiently expressed HSP90αE47A or HSP90β E42A. Using this approach, we identified a region in HSF1 that associates most strongly with
“closed” HSP90 (Supplemental Fig. 5). This motif is comprised of the HR-A/B trimerization domain in the context of a portion of the RD. Binding of “closed” HSP90 to this motif is
dramatically enhanced upon deletion of the HSF1 HR-C and TAD domains. Since the HR-C domain interacts with the HR-A/B domain to retard HSF1 trimerization under basal conditions, these data
are consistent with the preferential binding of “closed” HSP90 to HSF1 trimers. Residues in the C-terminal half of the HR-A/B domain (amino acids 183–214) are necessary and sufficient for
binding. Of note, while endogenous HSP90 shares the HR-A/B + RD binding motif, visualization of endogenous HSP90 binding requires deletion of HR-C and TAD domains, very likely due to the
weaker affinity of endogenous HSP90 (primarily existing in an “open” conformation) for client interactors. Taken together, these findings are not consistent with a role for HSP90 in
repressing HSF1 trimerization. In contrast to HSP90, endogenous HSP70 binds to the isolated RD as well as other domains in HSF1. Only the isolated HR-A/B and DBD domains display weak
interaction with HSP70. These data build upon earlier work by Shi _et al_. demonstrating repression of HSF1 transcriptional activity as a consequence of HSP70 binding to the TAD26,27. Our
observation that HSP70 also binds to the stress sensing RD may be relevant to the recent proposal that HSP70 provides an on/off switch for dynamic regulation of stress-induced HSF1
activation in yeast28. Whether HSP70 plays a similar role in regulating HSF1 activation in mammalian cells remains to be explored. We used these HSF1 truncation and deletion mutants to
identify the HR-A/B domain as necessary for dominant negative activity of HSF1 mutants lacking the TAD or the DBD. Importantly, and consistent with our model, we found that HSP90 binding to
the HR-A/B is not required for dominant-negative activity as it is not reversed by 17AAG. Instead, dominant-negative activity depends on the ability to form an intermolecular interaction
with other HSF1 monomers, suggesting that each monomer of an HSF1 trimer must be intact in order to attain optimal HSF1 transcriptional activity. A panel of clinically evaluated N-terminal
HSP90 inhibitors uniformly enhance HSF1 phosphorylation coincident with dissociation of HSF1 from HSP90. One or more drug-sensitive phosphorylation sites must lie outside of the RD, as an
HSF1 protein with all putative RD phosphorylation sites mutated to alanine (dPRD HSF1) still underwent a phosphorylation-dependent mobility shift upon HSP90 inhibitor treatment. We are
currently exploring the relevance of this observation to the activity of both wild type and dPRD HSF1, as well as the mechanistic impact of HSP90 inhibitor-mediated HSF1 phosphorylation on
its transcriptional activity. HSP90 ATP-competitive N-terminal inhibitors efficiently dissociate HSF1 from HSP90 “closed” mutants, likely as a consequence of ATP displacement and subsequent
disruption of N-domain dimerization40. If ATP-bound, transiently N-domain dimerized HSP90 (including stress-inducible HSP90α) provides feedback control of the HSR by removing HSF1 trimers
bound to heat shock promoter elements, HSP90 inhibition and subsequent dissociation from HSF1 would interfere with this process and would be predicted to prolong and/or intensify the HSR.
Consistent with this model, HSF1-CHiP data show that HSP90 inhibition alone and in combination with a brief heat shock significantly enhanced the duration of HSF1 binding to _HSP70_
promoter, compared to heat shock alone. When we assessed HSF1 transcriptional activity after these treatments (monitoring endogenous _HSP70_ mRNA levels by RT-qPCR), we found that
pre-exposure to HSP90 inhibitor markedly enhanced the degree and significantly extended the duration of _HSP70_ mRNA increase during recovery from a brief heat shock. HSP90 inhibition alone
caused a moderate increase in _HSP70_ transcription, likely due to disruption of the dynamic regulation of constitutive HSF1 activity in transformed cells, a phenomenon recently described by
Lindquist and colleagues11,12. In summary, this report increases our understanding of the molecular mechanisms that regulate HSF1 and the unique contribution of molecular chaperones,
including HSP90 and HSP70, to this process. While not precluding other possibilities, our findings are consistent with a role for HSP90 in attenuating the HSR in an ATP-dependent and
conformation-specific manner, a process that is antagonized by ATP-competitive HSP90 inhibitors. A role for HSP90 in disassembling transcriptional regulatory complexes has been described
previously45. Our data further suggest that, in contrast to HSP90, the readily observable interaction of endogenous HSP70 with HSF1, and our observation that the RD represents a major
interaction motif for this chaperone, support the possibility that HSP70 may provide constitutive, stress-sensitive repression of the HSR in mammalian cells. MATERIALS AND METHODS CELL
CULTURE HEK293 cells (American Type Culture Collection) were maintained in DMEM supplemented with 10% FBS. Cells were grown in 6-well or 96-well plates to 50% confluency and then transfected
for 16–24 hours with X-tremeGENE 9 transfection reagent (Roche) according to the provided protocol. HEK293 cells stably expressing an shRNA targeting HSF1 was obtained from Dr. Chengkai Dai
(Center for Cancer Research, NCI) and has been described previously46. REAGENTS 17AAG was obtained from the Developmental Therapeutics Program, NCI. STA9090 (ganetespib) and SNX2112 were
obtained from Synta Pharmaceuticals and Esanex, respectively. The C-terminal inhibitor KU32 was kindly provided by Dr. Brian Blagg (The University of Kansas, Lawrence, KS). MG132 and
molybdate were purchased from Sigma-Aldrich. All drugs were used at 10 μM for IP-WB or _hsp70_-reporter assays. PLASMIDS Sets of ATPase mutants of pcDNA-FLAG-HSP90α/β and pcDNA-HA-HSP90α/β
were made by the Quickchange method to alter designated amino acids. The series of C-terminal deletion constructs of pcDNA-FLAG-HSF1 were made by inserting a terminal stop codon into
designated regions by the Quickchange method. The series of N-terminal deletion constructs of pcDNA-FLAG-HSF1 were made by ligating truncated HSF1 insert with pcDNA-FLAG backbone by Gibson
assembly according to the provided protocol (New England Biolabs). IMMUNOPRECIPITATION AND WESTERN BLOTTING Cells were lysed with TGNET buffer [50 mM Tris HCl pH7.5, 5% Glycerol, 100 mM
NaCl, 2 mM EDTA, 0.5% Triton X-100] containing protease inhibitor cocktail and phosphatase inhibitor cocktail (Roche). Lysates were centrifuged at maximum speed for 15 minutes at 4 °C.
Supernatants were subjected to BCA protein assay for normalization, and then 40 μg of protein were used as input lysate to confirm protein expression. From remaining lysates, 700 μg of
protein were subjected to immunoprecipitation by addition of 40 μl of Anti-FLAG M2 affinity beads (Sigma-Aldrich, A2220) or HA affinity beads (clone HA-7, Sigma-Aldrich, A2095) followed by
incubation with rotation for 2 hours at 4 °C. After centrifugation, beads were washed 4 times with TGNET buffer. The proteins were then eluted with 30 μl of 2X SDS sample buffer by boiling
at 95 °C for 5 minutes. Subsequently, samples were subjected to SDS-PAGE followed by western blotting. The following antibodies were used for western blot: HSF1 (Santa Cruz, sc-13516), HSP90
(Enzo Life Science, ADI-SPA-835), HSP70 (Enzo Life Sciences, ADI-SPA-810), HA (Rockland, 600-401-384), and FLAG (Sigma-Aldrich, A8592). Additional Methods may be found in Supplemental
Information. DATA AVAILABILITY Materials and data generated in this study are available upon request to the corresponding authors (T.L.P. and L.N.). REFERENCES * Vihervaara, A. &
Sistonen, L. HSF1 at a glance. _Journal of cell science_ 127, 261–266, https://doi.org/10.1242/jcs.132605 (2014). Article CAS PubMed Google Scholar * Sorger, P. K. & Pelham, H. R.
Yeast heat shock factor is an essential DNA-binding protein that exhibits temperature-dependent phosphorylation. _Cell_ 54, 855–864 (1988). Article CAS PubMed Google Scholar * Perisic,
O., Xiao, H. & Lis, J. T. Stable binding of Drosophila heat shock factor to head-to-head and tail-to-tail repeats of a conserved 5 bp recognition unit. _Cell_ 59, 797–806 (1989). Article
CAS PubMed Google Scholar * Trinklein, N. D., Chen, W. C., Kingston, R. E. & Myers, R. M. Transcriptional regulation and binding of heat shock factor 1 and heat shock factor 2 to 32
human heat shock genes during thermal stress and differentiation. _Cell stress & chaperones_ 9, 21–28 (2004). Article CAS Google Scholar * Khaleque, M. A. _et al_. Heat shock factor
1 represses estrogen-dependent transcription through association with MTA1. _Oncogene_ 27, 1886–1893, https://doi.org/10.1038/sj.onc.1210834 (2008). Article CAS PubMed Google Scholar *
Mahat, D. B., Salamanca, H. H., Duarte, F. M., Danko, C. G. & Lis, J. T. Mammalian Heat Shock Response and Mechanisms Underlying Its Genome-wide Transcriptional Regulation. _Molecular
cell_ 62, 63–78, https://doi.org/10.1016/j.molcel.2016.02.025 (2016). Article CAS PubMed PubMed Central Google Scholar * Zou, J., Guo, Y., Guettouche, T., Smith, D. F. & Voellmy, R.
Repression of heat shock transcription factor HSF1 activation by HSP90 (HSP90 complex) that forms a stress-sensitive complex with HSF1. _Cell_ 94, 471–480 (1998). Article CAS PubMed
Google Scholar * Conde, R., Belak, Z. R., Nair, M., O’Carroll, R. F. & Ovsenek, N. Modulation of Hsf1 activity by novobiocin and geldanamycin. _Biochemistry and cell biology = Biochimie
et biologie cellulaire_ 87, 845–851, https://doi.org/10.1139/o09-049 (2009). Article CAS PubMed Google Scholar * Karagoz, G. E. & Rudiger, S. G. Hsp90 interaction with clients.
_Trends in biochemical sciences_ 40, 117–125, https://doi.org/10.1016/j.tibs.2014.12.002 (2015). Article CAS PubMed Google Scholar * Neef, D. W. _et al_. A direct regulatory interaction
between chaperonin TRiC and stress-responsive transcription factor HSF1. _Cell reports_ 9, 955–966, https://doi.org/10.1016/j.celrep.2014.09.056 (2014). Article CAS PubMed PubMed Central
Google Scholar * Mendillo, M. L. _et al_. HSF1 drives a transcriptional program distinct from heat shock to support highly malignant human cancers. _Cell_ 150, 549–562,
https://doi.org/10.1016/j.cell.2012.06.031 (2012). Article CAS PubMed PubMed Central Google Scholar * Santagata, S. _et al_. High levels of nuclear heat-shock factor 1 (HSF1) are
associated with poor prognosis in breast cancer. _Proceedings of the National Academy of Sciences of the United States of America_ 108, 18378–18383, https://doi.org/10.1073/pnas.1115031108
(2011). Article ADS CAS PubMed PubMed Central Google Scholar * Liao, Y. _et al_. Higher heat shock factor 1 expression in tumor stroma predicts poor prognosis in esophageal squamous
cell carcinoma patients. _Journal of translational medicine_ 13, 338, https://doi.org/10.1186/s12967-015-0703-x (2015). Article MathSciNet PubMed PubMed Central Google Scholar * Dai,
C., Whitesell, L., Rogers, A. B. & Lindquist, S. Heat shock factor 1 is a powerful multifaceted modifier of carcinogenesis. _Cell_ 130, 1005–1018,
https://doi.org/10.1016/j.cell.2007.07.020 (2007). Article CAS PubMed PubMed Central Google Scholar * Alarcon, S. V. _et al_. Tumor-intrinsic and tumor-extrinsic factors impacting
hsp90- targeted therapy. _Current molecular medicine_ 12, 1125–1141 (2012). Article CAS PubMed PubMed Central Google Scholar * Guettouche, T., Boellmann, F., Lane, W. S. & Voellmy,
R. Analysis of phosphorylation of human heat shock factor 1 in cells experiencing a stress. _BMC biochemistry_ 6, 4, https://doi.org/10.1186/1471-2091-6-4 (2005). Article PubMed PubMed
Central Google Scholar * Xu, Y. M., Huang, D. Y., Chiu, J. F. & Lau, A. T. Post-translational modification of human heat shock factors and their functions: a recent update by proteomic
approach. _Journal of proteome research_ 11, 2625–2634, https://doi.org/10.1021/pr201151a (2012). Article CAS PubMed Google Scholar * Littlefield, O. & Nelson, H. C. A new use for
the ‘wing’ of the ‘winged’ helix-turn-helix motif in the HSF-DNA cocrystal. _Nature structural biology_ 6, 464–470, https://doi.org/10.1038/8269 (1999). Article CAS PubMed Google Scholar
* Neudegger, T., Verghese, J., Hayer-Hartl, M., Hartl, F. U. & Bracher, A. Structure of human heat-shock transcription factor 1 in complex with DNA. _Nature structural & molecular
biology_ 23, 140–146, https://doi.org/10.1038/nsmb.3149 (2016). Article CAS Google Scholar * Schlessinger, A., Liu, J. & Rost, B. Natively unstructured loops differ from other loops.
_PLoS computational biology_ 3, e140, https://doi.org/10.1371/journal.pcbi.0030140 (2007). Article ADS PubMed PubMed Central Google Scholar * Rabindran, S. K., Haroun, R. I., Clos, J.,
Wisniewski, J. & Wu, C. Regulation of heat shock factor trimer formation: role of a conserved leucine zipper. _Science_ 259, 230–234 (1993). Article ADS CAS PubMed Google Scholar *
Newton, E. M., Knauf, U., Green, M. & Kingston, R. E. The regulatory domain of human heat shock factor 1 is sufficient to sense heat stress. _Molecular and cellular biology_ 16, 839–846
(1996). Article CAS PubMed PubMed Central Google Scholar * Budzynski, M. A., Puustinen, M. C., Joutsen, J. & Sistonen, L. Uncoupling Stress-Inducible Phosphorylation of Heat Shock
Factor 1 from Its Activation. _Molecular and cellular biology_ 35, 2530–2540, https://doi.org/10.1128/MCB.00816-14 (2015). Article CAS PubMed PubMed Central Google Scholar * Xie, Y.,
Chen, C., Stevenson, M. A., Auron, P. E. & Calderwood, S. K. Heat shock factor 1 represses transcription of the IL-1beta gene through physical interaction with the nuclear factor of
interleukin 6. _The Journal of biological chemistry_ 277, 11802–11810, https://doi.org/10.1074/jbc.M109296200 (2002). Article CAS PubMed Google Scholar * Hentze, N., Le Breton, L.,
Wiesner, J., Kempf, G. & Mayer, M. P. Molecular mechanism of thermosensory function of human heat shock transcription factor Hsf1. _eLife_ 5, https://doi.org/10.7554/eLife.11576 (2016).
* Shi, Y., Kroeger, P. E. & Morimoto, R. I. The carboxyl-terminal transactivation domain of heat shock factor 1 is negatively regulated and stress responsive. _Molecular and cellular
biology_ 15, 4309–4318 (1995). Article CAS PubMed PubMed Central Google Scholar * Shi, Y., Mosser, D. D. & Morimoto, R. I. Molecular chaperones as HSF1-specific transcriptional
repressors. _Genes & development_ 12, 654–666 (1998). Article CAS Google Scholar * Zheng, X. _et al_. Dynamic control of Hsf1 during heat shock by a chaperone switch and
phosphorylation. _eLife_ 5, https://doi.org/10.7554/eLife.18638 (2016). * Gidalevitz, T., Prahlad, V. & Morimoto, R. I. The stress of protein misfolding: from single cells to
multicellular organisms. _Cold Spring Harbor perspectives in biology_ 3, https://doi.org/10.1101/cshperspect.a009704 (2011). * Neckers, L. & Workman, P. Hsp90 molecular chaperone
inhibitors: are we there yet? _Clinical cancer research: an official journal of the American Association for Cancer Research_ 18, 64–76, https://doi.org/10.1158/1078-0432.CCR-11-1000 (2012).
Article CAS Google Scholar * Trepel, J., Mollapour, M., Giaccone, G. & Neckers, L. Targeting the dynamic HSP90 complex in cancer. _Nature reviews. Cancer_ 10, 537–549,
https://doi.org/10.1038/nrc2887 (2010). Article CAS PubMed Google Scholar * Barrott, J. J. & Haystead, T. A. Hsp90, an unlikely ally in the war on cancer. _The FEBS journal_ 280,
1381–1396, https://doi.org/10.1111/febs.12147 (2013). Article CAS PubMed Google Scholar * Johnson, M. L. _et al_. Phase I/II Study of HSP90 Inhibitor AUY922 and Erlotinib for EGFR-Mutant
Lung Cancer With Acquired Resistance to Epidermal Growth Factor Receptor Tyrosine Kinase Inhibitors. _Journal of clinical oncology: official journal of the American Society of Clinical
Oncology_ 33, 1666–1673, https://doi.org/10.1200/JCO.2014.59.7328 (2015). Article CAS Google Scholar * Do, K. _et al_. Phase I study of the heat shock protein 90 (Hsp90) inhibitor
onalespib (AT13387) administered on a daily for 2 consecutive days per week dosing schedule in patients with advanced solid tumors. _Investigational new drugs_ 33, 921–930,
https://doi.org/10.1007/s10637-015-0255-1 (2015). Article CAS PubMed Google Scholar * Shapiro, G. I. _et al_. First-in-human phase I dose escalation study of a second-generation
non-ansamycin HSP90 inhibitor, AT13387, in patients with advanced solid tumors. _Clinical cancer research: an official journal of the American Association for Cancer Research_ 21, 87–97,
https://doi.org/10.1158/1078-0432.CCR-14-0979 (2015). Article CAS Google Scholar * Prince, T. L. _et al_. Client Proteins and Small Molecule Inhibitors Display Distinct Binding
Preferences for Constitutive and Stress-Induced HSP90 Isoforms and Their Conformationally Restricted Mutants. _PloS one_ 10, e0141786, https://doi.org/10.1371/journal.pone.0141786 (2015).
Article PubMed PubMed Central Google Scholar * Southworth, D. R. & Agard, D. A. Species-dependent ensembles of conserved conformational states define the Hsp90 chaperone ATPase
cycle. _Molecular cell_ 32, 631–640, https://doi.org/10.1016/j.molcel.2008.10.024 (2008). Article CAS PubMed PubMed Central Google Scholar * Hartson, S. D., Thulasiraman, V., Huang, W.,
Whitesell, L. & Matts, R. L. Molybdate inhibitshsp90, induces structural changes in its C-terminal domain, and alters its interactions with substrates. _Biochemistry_ 38, 3837–3849,
https://doi.org/10.1021/bi983027s (1999). Article CAS PubMed Google Scholar * Taipale, M. _et al_. Quantitative analysis of HSP90-client interactions reveals principles of substrate
recognition. _Cell_ 150, 987–1001, https://doi.org/10.1016/j.cell.2012.06.047 (2012). Article CAS PubMed PubMed Central Google Scholar * Beebe, K. _et al_. Posttranslational
modification and conformational state of heat shock protein 90 differentially affect binding of chemically diverse small molecule inhibitors. _Oncotarget_ 4, 1065–1074,
https://doi.org/10.18632/oncotarget.1099 (2013). Article PubMed PubMed Central Google Scholar * Donnelly, A. & Blagg, B. S. Novobiocin and additional inhibitors of the Hsp90
C-terminal nucleotide-binding pocket. _Current medicinal chemistry_ 15, 2702–2717 (2008). Article CAS PubMed PubMed Central Google Scholar * Kusuma, B. R. _et al_. Synthesis and
evaluation of novologues as C-terminal Hsp90 inhibitors with cytoprotective activity against sensory neuron glucotoxicity. _Journal of medicinal chemistry_ 55, 5797–5812,
https://doi.org/10.1021/jm300544c (2012). Article CAS PubMed PubMed Central Google Scholar * Anckar, J. & Sistonen, L. Regulation of HSF1 function in the heat stress response:
implications in aging and disease. _Annual review of biochemistry_ 80, 1089–1115, https://doi.org/10.1146/annurev-biochem-060809-095203 (2011). Article CAS PubMed Google Scholar * Ali,
A., Bharadwaj, S., O’Carroll, R. & Ovsenek, N. HSP90 interacts with and regulates the activity of heat shock factor 1 in Xenopus oocytes. _Molecular and cellular biology_ 18, 4949–4960
(1998). Article CAS PubMed PubMed Central Google Scholar * Freeman, B. C. & Yamamoto, K. R. Disassembly of transcriptional regulatory complexes by molecular chaperones. _Science_
296, 2232–2235, https://doi.org/10.1126/science.1073051 (2002). Article ADS CAS PubMed Google Scholar * Tang, Z. _et al_. MEK guards proteome stability and inhibits tumor-suppressive
amyloidogenesis via HSF1. _Cell_ 160, 729–744, https://doi.org/10.1016/j.cell.2015.01.028 (2015). Article CAS PubMed PubMed Central Google Scholar Download references ACKNOWLEDGEMENTS
This study was supported by funds from the Intramural Research Program, National Cancer Institute, Center for Cancer Research (L.N.). T.K. was supported by a JSPS Research Fellowship for
Japanese Biomedical and Behavioral Researchers at NIH. We dedicate this paper to the memory of Dr. Susan Lindquist, a friend and a pioneer in the study of molecular chaperones. AUTHOR
INFORMATION Author notes * Toshiki Kijima, Thomas L. Prince and Megan L. Tigue contributed equally to this work. AUTHORS AND AFFILIATIONS * Urologic Oncology Branch, Center for Cancer
Research, National Cancer Institute, National Institutes of Health, Bethesda, MD, 20892, United States Toshiki Kijima, Thomas L. Prince, Megan L. Tigue, Kendrick H. Yim, Harvey Schwartz,
Kristin Beebe & Len Neckers * Urology Department, Geisinger Clinic, WCR 221, Danville, PA, 17821, United States Thomas L. Prince & Heinric Williams * Developmental Therapeutics
Branch, Center for Cancer Research, National Cancer Institute, National Institutes of Health, Bethesda, MD, 20892, United States Sunmin Lee & Jane B. Trepel * Faculty of Science and
Engineering, Cell Biology, Åbo Akademi University, BioCity, Tykistökatu 6, Turku, FIN-20520, Finland Marek A. Budzynski & Lea Sistonen * Radiation Oncology, Harvard Medical School, Beth
Israel Deaconess Medical Center, 330 Brookline Ave., Boston, MA, 02215, United States Stuart Calderwood Authors * Toshiki Kijima View author publications You can also search for this author
inPubMed Google Scholar * Thomas L. Prince View author publications You can also search for this author inPubMed Google Scholar * Megan L. Tigue View author publications You can also search
for this author inPubMed Google Scholar * Kendrick H. Yim View author publications You can also search for this author inPubMed Google Scholar * Harvey Schwartz View author publications You
can also search for this author inPubMed Google Scholar * Kristin Beebe View author publications You can also search for this author inPubMed Google Scholar * Sunmin Lee View author
publications You can also search for this author inPubMed Google Scholar * Marek A. Budzynski View author publications You can also search for this author inPubMed Google Scholar * Heinric
Williams View author publications You can also search for this author inPubMed Google Scholar * Jane B. Trepel View author publications You can also search for this author inPubMed Google
Scholar * Lea Sistonen View author publications You can also search for this author inPubMed Google Scholar * Stuart Calderwood View author publications You can also search for this author
inPubMed Google Scholar * Len Neckers View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS T.K. designed experiments, performed experiments and
helped write the paper; T.L.P. designed experiments, performed experiments, interpreted the data and helped write the paper; M.L.T., K.H.Y., H.S., K.B. performed experiments and interpreted
the data; S.L. and J.B.T. performed experiments, interpreted the data and helped write the paper; M.A.B. and L.S. provided reagents, interpreted data and helped write the paper; H.W. and
S.C. interpreted data and helped write the paper; L.N. oversaw the whole project, devised experiments, interpreted data and wrote the paper. CORRESPONDING AUTHORS Correspondence to Thomas L.
Prince or Len Neckers. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. ADDITIONAL INFORMATION PUBLISHER'S NOTE: Springer Nature remains neutral with
regard to jurisdictional claims in published maps and institutional affiliations. ELECTRONIC SUPPLEMENTARY MATERIAL SUPPLEMENTARY INFORMATION RIGHTS AND PERMISSIONS OPEN ACCESS This article
is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you
give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material
in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative
Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a
copy of this license, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Kijima, T., Prince, T.L., Tigue, M.L. _et al._ HSP90
inhibitors disrupt a transient HSP90-HSF1 interaction and identify a noncanonical model of HSP90-mediated HSF1 regulation. _Sci Rep_ 8, 6976 (2018).
https://doi.org/10.1038/s41598-018-25404-w Download citation * Received: 08 November 2017 * Accepted: 16 April 2018 * Published: 03 May 2018 * DOI: https://doi.org/10.1038/s41598-018-25404-w
SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy
to clipboard Provided by the Springer Nature SharedIt content-sharing initiative