- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT G-protein-coupled receptors (GPCRs) mediate many critical physiological processes. Their spatial organization in plasma membrane (PM) domains is believed to encode signaling
specificity and efficiency. However, the existence of domains and, crucially, the mechanism of formation of such putative domains remain elusive. Here, live-cell imaging (corrected for
topography-induced imaging artifacts) conclusively established the existence of PM domains for GPCRs. Paradoxically, energetic coupling to extremely shallow PM curvature (<1 µm−1) emerged
as the dominant, necessary and sufficient molecular mechanism of GPCR spatiotemporal organization. Experiments with different GPCRs, H-Ras, Piezo1 and epidermal growth factor receptor,
suggest that the mechanism is general, yet protein specific, and can be regulated by ligands. These findings delineate a new spatiomechanical molecular mechanism that can transduce to
domain-based signaling any mechanical or chemical stimulus that affects the morphology of the PM and suggest innovative therapeutic strategies targeting cellular shape. Access through your
institution Buy or subscribe This is a preview of subscription content, access via your institution ACCESS OPTIONS Access through your institution Access Nature and 54 other Nature Portfolio
journals Get Nature+, our best-value online-access subscription $29.99 / 30 days cancel any time Learn more Subscribe to this journal Receive 12 print issues and online access $259.00 per
year only $21.58 per issue Learn more Buy this article * Purchase on SpringerLink * Instant access to full article PDF Buy now Prices may be subject to local taxes which are calculated
during checkout ADDITIONAL ACCESS OPTIONS: * Log in * Learn about institutional subscriptions * Read our FAQs * Contact customer support SIMILAR CONTENT BEING VIEWED BY OTHERS THE SPATIAL
DISTRIBUTION OF GPCR AND GΒΓ ACTIVITY ACROSS A CELL DICTATES PIP3 DYNAMICS Article Open access 16 February 2023 MEMBRANE CURVATURE GOVERNS THE DISTRIBUTION OF PIEZO1 IN LIVE CELLS Article
Open access 03 December 2022 ENDOSOME POSITIONING COORDINATES SPATIALLY SELECTIVE GPCR SIGNALING Article 27 July 2023 DATA AVAILABILITY The structure of the inactive β1AR is available from
the Protein Data Bank with accession code 2YCW. Source data are provided with this paper. CODE AVAILABILITY Algorithms used by custom analysis code are described in detail in the Methods.
Code is available upon reasonable request. REFERENCES * Pierce, K. L., Premont, R. T. & Lefkowitz, R. J. Seven-transmembrane receptors. _Nat. Rev. Mol. Cell Biol._ 3, 639–650 (2002).
Article CAS PubMed Google Scholar * Weis, W. I. & Kobilka, B. K. The molecular bases of G protein-coupled receptor activation. _Annu. Rev. Biochem._ 87, 879–919 (2018). Article
Google Scholar * Wootten, D., Christopoulos, A., Marti-Solano, M., Babu, M. M. & Sexton, P. M. Mechanisms of signalling and biased agonism in G protein-coupled receptors. _Nat. Rev.
Mol. Cell Biol._ 19, 638–653 (2018). Article CAS PubMed Google Scholar * Nikolaev, V. O. et al. β2-Adrenergic receptor redistribution in heart failure changes cAMP compartmentation.
_Science_ 327, 1653–1657 (2010). Article CAS PubMed Google Scholar * Irannejad, R. et al. Functional selectivity of GPCR-directed drug action through location bias. _Nat. Chem. Biol._
13, 799–806 (2017). Article CAS PubMed PubMed Central Google Scholar * Briddon, S. J., Kilpatrick, L. E. & Hill, S. J. Studying GPCR pharmacology in membrane microdomains:
fluorescence correlation spectroscopy comes of age. _Trends Pharmacol. Sci._ 39, 158–174 (2018). Article CAS PubMed Google Scholar * Insel, P. A. et al. Compartmentation of
G-protein-coupled receptors and their signalling components in lipid rafts and caveolae. _Biochem. Soc. Trans._ 33, 1131-4 (2005). Article PubMed Google Scholar * Pontier, S. M. et al.
Cholesterol-dependent separation of the β2-adrenergic receptor from its partners determines signaling efficacy: insight into nanoscale organization of signal transduction. _J. Biol. Chem._
283, 24659–24672 (2008). Article CAS PubMed PubMed Central Google Scholar * Lohse, M. J. & Hofmann, K. P. Spatial and temporal aspects of signaling by G-protein-coupled receptors.
_Mol. Pharmacol._ 88, 572–578 (2015). Article CAS PubMed Google Scholar * Calebiro, D. & Sungkaworn, T. Single-molecule imaging of GPCR interactions. _Trends Pharmacol. Sci._ 39,
109–122 (2018). Article CAS PubMed Google Scholar * Halls, M. L. et al. Plasma membrane localization of the μ-opioid receptor controls spatiotemporal signaling. _Sci. Signal._ 9, ra16
(2016). Article PubMed Google Scholar * Sungkaworn, T. et al. Single-molecule imaging reveals receptor-G protein interactions at cell surface hot spots. _Nature_ 550, 543–547 (2017).
Article CAS PubMed Google Scholar * Sezgin, E., Levental, I., Mayor, S. & Eggeling, C. The mystery of membrane organization: composition, regulation and roles of lipid rafts. _Nat.
Rev. Mol. Cell Biol._ 18, 361–374 (2017). Article CAS PubMed PubMed Central Google Scholar * Annibale, P. & Lohse, M. J. Spatial heterogeneity in molecular brightness. _Nat.
Methods_ 17, 273–275 (2020). Article CAS PubMed Google Scholar * Adler, J., Shevchuk, A. I., Novak, P., Korchev, Y. E. & Parmryd, I. Plasma membrane topography and interpretation of
single-particle tracks. _Nat. Methods_ 7, 170–171 (2010). Article CAS PubMed Google Scholar * Eichel, K. & von Zastrow, M. Subcellular organization of GPCR signaling. _Trends
Pharmacol. Sci._ 39, 200–208 (2018). Article CAS PubMed PubMed Central Google Scholar * Rosholm, K. R. et al. Membrane curvature regulates ligand-specific membrane sorting of GPCRs in
living cells. _Nat. Chem. Biol._ 13, 724–729 (2017). Article CAS PubMed Google Scholar * Paszek, M. J. et al. Scanning angle interference microscopy reveals cell dynamics at the
nanoscale. _Nat. Methods_ 9, 825–827 (2012). Article CAS PubMed PubMed Central Google Scholar * Elliott, H. et al. Myosin II controls cellular branching morphogenesis and migration in
three dimensions by minimizing cell-surface curvature. _Nat. Cell Biol._ 17, 137–147 (2015). Article CAS PubMed PubMed Central Google Scholar * Kabbani, A. M. & Kelly, C. V. The
detection of nanoscale membrane bending with polarized localization microscopy. _Biophys. J._ 113, 1782–1794 (2017). Article CAS PubMed PubMed Central Google Scholar * van Rheenen, J.
& Jalink, K. Agonist-induced PIP2 hydrolysis inhibits cortical actin dynamics: regulation at a global but not at a micrometer scale. _Mol. Biol. Cell_ 13, 3257–3267 (2002). Article
PubMed PubMed Central Google Scholar * Clack, N. G. & Groves, J. T. Many-particle tracking with nanometer resolution in three dimensions by reflection interference contrast
microscopy. _Langmuir_ 21, 6430–6435 (2005). Article CAS PubMed Google Scholar * Calebiro, D. et al. Single-molecule analysis of fluorescently labeled G-protein-coupled receptors reveals
complexes with distinct dynamics and organization. _Proc. Natl Acad. Sci. USA_ 110, 743–748 (2013). Article CAS PubMed Google Scholar * May, S. & Ben-Shaul, A. A molecular model for
lipid-mediated interaction between proteins in membranes. _Phys. Chem. Chem. Phys._ 2, 4494–4502 (2000). Article CAS Google Scholar * Sperotto, M. M., May, S. & Baumgaertner, A.
Modelling of proteins in membranes. _Chem. Phys. Lipids_ 141, 2–29 (2006). Article CAS PubMed Google Scholar * Larsen, J. B. et al. Membrane curvature enables N-Ras lipid anchor sorting
to liquid-ordered membrane phases. _Nat. Chem. Biol._ 11, 192–194 (2015). Article CAS PubMed Google Scholar * Larsen, J. B. et al. Membrane curvature and lipid composition synergize to
regulate N-Ras anchor recruitment. _Biophys. J._ 113, 1269–1279 (2017). Article CAS PubMed PubMed Central Google Scholar * Larsen, J. B. et al. How membrane geometry regulates protein
sorting independently of mean curvature. _ACS Cent. Sci._ 6, 1159–1168 (2020). Article CAS PubMed PubMed Central Google Scholar * Moukhametzianov, R. et al. Two distinct conformations
of helix 6 observed in antagonist-bound structures of a β1-adrenergic receptor. _Proc. Natl Acad. Sci. USA_ 108, 8228–8232 (2011). Article CAS PubMed PubMed Central Google Scholar *
Lorent, J. H. et al. Plasma membranes are asymmetric in lipid unsaturation, packing and protein shape. _Nat. Chem. Biol._ 16, 644–652 (2020). Article CAS PubMed PubMed Central Google
Scholar * Marrink, S. J. et al. Computational modeling of realistic cell membranes. _Chem. Rev._ 119, 6184–6226 (2019). Article CAS PubMed PubMed Central Google Scholar * Latorraca, N.
R., Venkatakrishnan, A. J. & Dror, R. O. GPCR dynamics: structures in motion. _Chem. Rev._ 117, 139–155 (2017). Article CAS PubMed Google Scholar * Zhao, W. et al. Nanoscale
manipulation of membrane curvature for probing endocytosis in live cells. _Nat. Nanotechnol._ 12, 750–756 (2017). Article CAS PubMed PubMed Central Google Scholar * McMahon, H. T. &
Gallop, J. L. Membrane curvature and mechanisms of dynamic cell membrane remodelling. _Nature_ 438, 590–596 (2005). Article CAS PubMed Google Scholar * Aimon, S. et al. Membrane shape
modulates transmembrane protein distribution. _Dev. Cell_ 28, 212–218 (2014). Article CAS PubMed PubMed Central Google Scholar * Brown, M. F. Curvature forces in membrane lipid–protein
interactions. _Biochemistry_ 51, 9782–9795 (2012). Article CAS PubMed Google Scholar * Hossein, A. & Deserno, M. Spontaneous curvature, differential stress, and bending modulus of
asymmetric lipid membranes. _Biophys. J._ 118, 624–642 (2020). Article CAS PubMed Google Scholar * Soubias, O., Teague, W. E. Jr, Hines, K. G. & Gawrisch, K. Rhodopsin/lipid
hydrophobic matching-rhodopsin oligomerization and function. _Biophys. J._ 108, 1125–1132 (2015). Article CAS PubMed PubMed Central Google Scholar * Joyce, G., Robertson, B. D. &
Williams, K. J. A modified agar pad method for mycobacterial live-cell imaging. _BMC Res. Notes_ 4, 73 (2011). * Bathe-Peters, M. et al. Visualization of β-adrenergic receptor dynamics and
differential localization in cardiomyocytes. _Proc. Natl Acad. Sci. USA_ 118, e2101119118 (2021). Article CAS PubMed PubMed Central Google Scholar * Zhang, Y. et al. Cryo-EM structure
of the activated GLP-1 receptor in complex with a G protein. _Nature_ 546, 248–253 (2017). Article CAS PubMed PubMed Central Google Scholar * Murthy, S. E., Dubin, A. E. &
Patapoutian, A. Piezos thrive under pressure: mechanically activated ion channels in health and disease. _Nat. Rev. Mol. Cell Biol._ 18, 771–783 (2017). Article CAS PubMed Google Scholar
* Zhou, R., Han, B., Xia, C. & Zhuang, X. Membrane-associated periodic skeleton is a signaling platform for RTK transactivation. _Science_ 365, 929–934 (2019). Article CAS PubMed
PubMed Central Google Scholar * Antonny, B. Mechanisms of membrane curvature sensing. _Annu. Rev. Biochem._ 80, 101–123 (2011). Article CAS PubMed Google Scholar * Iversen, L.,
Mathiasen, S., Larsen, J. B. & Stamou, D. Membrane curvature bends the laws of physics and chemistry. _Nat. Chem. Biol._ 11, 822–825 (2015). Article CAS PubMed Google Scholar *
Bassereau, P. et al. The 2018 biomembrane curvature and remodeling roadmap. _J. Phys. D Appl. Phys._ 51, 343001 (2018). Article PubMed PubMed Central Google Scholar * Lingwood, D. &
Simons, K. Lipid rafts as a membrane-organizing principle. _Science_ 327, 46–50 (2010). Article CAS PubMed Google Scholar * Hatzakis, N. S. et al. How curved membranes recruit
amphipathic helices and protein anchoring motifs. _Nat. Chem. Biol._ 5, 835–841 (2009). Article CAS PubMed Google Scholar * Huang, W. et al. Structure of the neurotensin receptor 1 in
complex with β-arrestin 1. _Nature_ 579, 303–308 (2020). Article CAS PubMed PubMed Central Google Scholar * Foerster, F. et al. Targeting the actin cytoskeleton: selective antitumor
action via trapping PKCε. _Cell Death Dis._ 5, e1398 (2014). Article CAS PubMed PubMed Central Google Scholar * Roffay, C. et al. Passive coupling of membrane tension and cell volume
during active response of cells to osmosis. _Proc. Natl Acad. Sci. USA_ 118, e2103228118 (2021). Article CAS PubMed PubMed Central Google Scholar * Zimmerberg, J. & Kozlov, M. M.
How proteins produce cellular membrane curvature. _Nat. Rev. Mol. Cell Biol._ 7, 9–19 (2006). Article CAS PubMed Google Scholar * Wolfenson, H., Yang, B. & Sheetz, M. Steps in
mechanotransduction pathways that control cell morphology. _Annu. Rev. Physiol._ 81, 585–605 (2019). Article CAS PubMed Google Scholar * Zimmerberg, J. & Gawrisch, K. The physical
chemistry of biological membranes. _Nat. Chem. Biol._ 2, 564–567 (2006). Article CAS PubMed Google Scholar * Staudt, T. et al. Far-field optical nanoscopy with reduced number of state
transition cycles. _Opt. Express_ 19, 5644–5657 (2011). Article PubMed Google Scholar * Shelton, E., Serwane, F. & Campas, O. Geometrical characterization of fluorescently labelled
surfaces from noisy 3D microscopy data. _J. Microsc._ 269, 259–268 (2018). Article CAS PubMed Google Scholar * Lohse, M. J., Nuber, S. & Hoffmann, C. Fluorescence/bioluminescence
resonance energy transfer techniques to study G-protein-coupled receptor activation and signaling. _Pharmacol. Rev._ 64, 299–336 (2012). Article CAS PubMed Google Scholar * Lambert, N.
A. & Javitch, J. A. CrossTalk opposing view: weighing the evidence for class A GPCR dimers, the jury is still out. _J. Physiol._ 592, 2443–2445 (2014). Article CAS PubMed PubMed
Central Google Scholar * Barbotin, A. et al. z-STED imaging and spectroscopy to investigate nanoscale membrane structure and dynamics. _Biophys. J._ 118, 2448–2457 (2020). Article CAS
PubMed PubMed Central Google Scholar * Uline, M. J. & Szleifer, I. Mode specific elastic constants for the gel, liquid-ordered, and liquid-disordered phases of DPPC/DOPC/cholesterol
model lipid bilayers. _Faraday Discuss._ 161, 177–191 (2013). Article CAS PubMed PubMed Central Google Scholar * Keppler, A. et al. A general method for the covalent labeling of fusion
proteins with small molecules in vivo. _Nat. Biotechnol._ 21, 86–89 (2003). Article CAS PubMed Google Scholar * Sungkaworn, T., Rieken, F., Lohse, M. J. & Calebiro, D.
High-resolution spatiotemporal analysis of receptor dynamics by single-molecule fluorescence microscopy. _J. Vis. Exp_. https://doi.org/10.3791/51784 (2014). * Maiellaro, I., Lohse, M. J.,
Kittel, R. J. & Calebiro, D. Real-time monitoring of GPCR/cAMP signalling by FRET and single-molecule microscopy. _Horm. Metab. Res._ 46, 827–832 (2014). Article PubMed Google Scholar
* Curtis, A. The mechanism of adhesion of cell to glass: a study by Interference reflection microscopy. _J. Cell Biol._ 20, 199–215 (1964). Article CAS PubMed PubMed Central Google
Scholar * Limozin, L. & Sengupta, K. Quantitative reflection interference contrast microscopy (RICM) in soft matter and cell adhesion. _ChemPhysChem_ 10, 2752–2768 (2009). Article CAS
PubMed Google Scholar * Barr, V. A. & Bunnell, S. C. Interference reflection microscopy. _Curr. Protoc. Cell Biol._ 4, 4.23 (2009). * Kukulski, W. et al. Precise, correlated
fluorescence microscopy and electron tomography of lowicryl sections using fluorescent fiducial markers. _Methods Cell. Biol._ 111, 235–257 (2012). Article CAS PubMed Google Scholar *
Biswas, A., Alex, A. & Sinha, B. Mapping cell membrane fluctuations reveals their active regulation and transient heterogeneities. _Biophys. J._ 113, 1768–1781 (2017). Article CAS
PubMed PubMed Central Google Scholar * Vicidomini, G. et al. STED nanoscopy with time-gated detection: theoretical and experimental aspects. _PLoS ONE_ 8, e54421 (2013). Article CAS
PubMed PubMed Central Google Scholar * Manders, E. M. M., Verbeek, F. J. & Aten, J. A. Measurement of co-localization of objects in dual-colour confocal images. _J. Microsc._ 169,
375–382 (1993). Article CAS PubMed Google Scholar Download references ACKNOWLEDGEMENTS This work was supported by the Novo Nordisk Foundation (grant NNF17OC0028176; D.S.) and NIGMS R35
GM118167 (O.D.W.). We thank S. Hell, C. Eggeling and E. Sezgin for initial assistance with 3D STED and J. T. Groves, R. Dimova, D. Wustner and K. Madsen for helpful discussions on the
project. We are also thankful to P. Lappalainen for the GFP–actin plasmid, P. J. Verveer for the EGFR–SNAP plasmid, N. Schmitt for HL-1 cells and K. L. Madsen for COS-7 cells. We thank T.
Izard for pcDNA4-GFP-vinculin (Addgene plasmid 107153). We are grateful to D. Gadella for mNeonGreen–Rab7 (Addgene plasmid 129603), 4xmts-mNeonGreen (Addgene plasmid 98876) and
lysozyme(1–31)-KDEL–mNeonGreen (Addgene plasmid 137804). We thank Y. Schwab and A. Kolovou for their help and expertise in cryo-EM. AUTHOR INFORMATION Author notes * These authors
contributed equally: Gabriele Kockelkoren, Line Lauritsen. AUTHORS AND AFFILIATIONS * Center for Geometrically Engineered Cellular Membranes, Department of Chemistry, University of
Copenhagen, Copenhagen, Denmark Gabriele Kockelkoren, Line Lauritsen, Christopher G. Shuttle, Eleftheria Kazepidou, Ivana Vonkova, Artù Breuer, Mark Uline & Dimitrios Stamou * Howard
Hughes Medical Institute, Department of Neuroscience, Dorris Neuroscience Center, Scripps Research, La Jolla, CA, USA Yunxiao Zhang * Department of Chemical Engineering, Biomedical
Engineering Program, University of South Carolina, Columbia, SC, USA Celeste Kennard & Mark Uline * Cardiovascular Research Institute, University of California, San Francisco, CA, USA
Rachel M. Brunetti & Orion D. Weiner * Department of Biochemistry and Biophysics, University of California, San Francisco, CA, USA Rachel M. Brunetti & Orion D. Weiner * Center for
Geometrically Engineered Cellular Membranes, University of California, San Francisco, CA, USA Rachel M. Brunetti & Orion D. Weiner * Max-Planck-Institute for Medical Research, Optical
Microscopy Facility, Heidelberg, Germany Elisa D’Este * Atomos Biotech, Copenhagen, Denmark Dimitrios Stamou Authors * Gabriele Kockelkoren View author publications You can also search for
this author inPubMed Google Scholar * Line Lauritsen View author publications You can also search for this author inPubMed Google Scholar * Christopher G. Shuttle View author publications
You can also search for this author inPubMed Google Scholar * Eleftheria Kazepidou View author publications You can also search for this author inPubMed Google Scholar * Ivana Vonkova View
author publications You can also search for this author inPubMed Google Scholar * Yunxiao Zhang View author publications You can also search for this author inPubMed Google Scholar * Artù
Breuer View author publications You can also search for this author inPubMed Google Scholar * Celeste Kennard View author publications You can also search for this author inPubMed Google
Scholar * Rachel M. Brunetti View author publications You can also search for this author inPubMed Google Scholar * Elisa D’Este View author publications You can also search for this author
inPubMed Google Scholar * Orion D. Weiner View author publications You can also search for this author inPubMed Google Scholar * Mark Uline View author publications You can also search for
this author inPubMed Google Scholar * Dimitrios Stamou View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS D.S. conceived the strategy and was
responsible for project management and supervision. L.L. developed and validated the principle of 3D imaging with help from C.G.S. and G.K. G.K. developed and validated the cell compression
assay with help from R.M.B. and O.D.W. G.K. and D.S. designed the experiments. G.K. collected data with help from L.L. G.K. analyzed data with help from L.L. M.U. performed all theoretical
calculations. C.G.S. was the principal software developer in the project. I.V. performed thin-section cryo-EM experiments. Y.Z. made constructs and provided help with Piezo1 experiments.
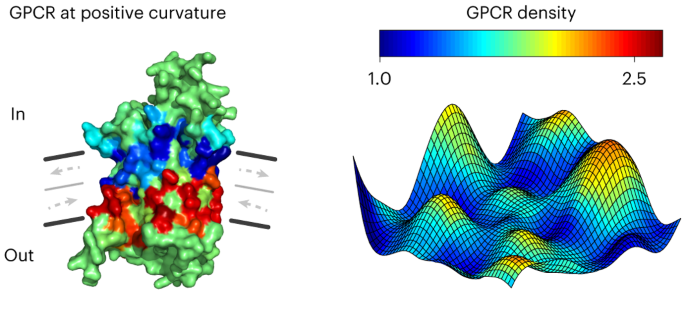
C.K. developed visualization of color-coded amino acid residues onto the receptor structure in Fig. 2a,b, A.B. and E.K. contributed to conceptualization and preliminary experiments. E.D.
provided help with STED microscopy. D.S. and G.K. wrote the main text. G.K. prepared all main and Supplementary figures with the help of L.L. for several Supplementary figures. M.U. and L.L.
contributed to the Supplementary Information. All authors discussed the results and commented on the manuscript. CORRESPONDING AUTHORS Correspondence to Mark Uline or Dimitrios Stamou.
ETHICS DECLARATIONS COMPETING INTERESTS D.S. is the founder of Atomos Biotech. The other authors declare no competing interests. PEER REVIEW PEER REVIEW INFORMATION _Nature Chemical Biology_
thanks Paolo Annibale, Juan Vanegas, and Wade Zeno for their contribution to the peer review of this work. ADDITIONAL INFORMATION PUBLISHER’S NOTE Springer Nature remains neutral with
regard to jurisdictional claims in published maps and institutional affiliations. EXTENDED DATA EXTENDED DATA FIG. 1 2D IMAGING REVEALS SPATIAL VARIATIONS IN RECEPTOR INTENSITY THAT CANNOT
BE INTERPRETED WITHOUT KNOWLEDGE OF MEMBRANE TOPOGRAPHY. (A) Cartoons represent slices of membranes imaged by Confocal or total internal fluorescence microscopy (TIRF) microscopy at the cell
equator and at the plasma membrane. (B-F) Confocal (b-d) or TIRF (e-f) images of the PM of HEK293 labeled with CellMask and/or the β1AR. All images are recorded at the basolateral membrane
except for (B, left) which is at the cell equator. The heterogenous spatial distribution of _intensity_ (domains of high/low intensity) cannot be interpreted as variations in _density_
without prior knowledge of membrane topography. Color scales show relative intensity that is the smallest intensity present in an image is set to black. Data is from _n__R_ = 3. Scalebars:
(a-b) 5 µm; (c-f) 500 nm. EXTENDED DATA FIG. 2 QUANTITATIVE MEASUREMENT OF PROTEIN DENSITY AND DIFFUSION REQUIRES IMAGING OF THE PLASMA MEMBRANE IN 3D TO CORRECT TOPOGRAPHY-INDUCED ARTIFACTS
THAT MIMIC THE APPEARANCE OF DOMAINS IN 2D PROJECTIONS. (A) Experimentally obtained 3D membrane topography map of the adherent plasma membrane of a HEK293 cell labelled using CellMask. (B)
Illustration of the confocal excitation volume approximated by an ellipsoid (blue) and the tangent plane at a given point of the membrane (green). The membrane tilt angle θ (angle between
the tangent plane and the imaging plane) varies between 0° – 90°. (C) The cross-sectional area between the plane and the ellipsoid (approximated by a cylinder for simplicity) scales with sec
_θ_ and serves as an approximation of the tilted membrane area. (D) An experimentally obtained 3D membrane topography map to which we computationally assigned a uniform receptor density.
(E) Because of the variations in membrane topography, the 2D density projection of the uniform surface in (d) erroneously suggests the existence of GPCR domains. (F) For similar reasons, the
spatially homogeneous diffusion in the 3D surface shown in (d) will erroneously appear to be heterogeneous if projected in 2D. (G) Schematic illustrations. The 2D projection of a uniformly
labelled membrane of varying topography (left), can erroneously produce the appearance of 2D domains that cannot a priori be distinguished from bona fide variations in membrane label density
(right). Source data EXTENDED DATA FIG. 3 GENERATION OF HIGH-ACCURACY TOPOGRAPHY MAPS OF PLASMA MEMBRANES OF LIVING CELLS. (A, B) Fluorescence confocal image of HEK293 cells over-expressing
SNAP-β1AR (a) and CellMask (b). Data β1AR is representative for _n__R_ = 4 and CellMask _n__R_ = 3 replicates. (C, D) Illustration of an XZY-stack (dz/dx/dy = 30 nm) of the highlighted area
in (a) and (b). (E, F) Extracted intensity Z-profiles of linescans highlighted in (c) and (d) and their corresponding Gaussian fits overlaid. The axial position of the Gaussian peak
corresponds to the Z position of the membrane, whereas the amplitude of the Gaussian peak is proportional to protein or Cellmask density. (G) Topography map of the area shown in (b)
reconstructed from the Z positions obtained from the Gaussian fitting. Color scale represents membrane height in nm. (H) Local error weighted quadric fit for a 3 × 3 pixel, 90 nm x 90 nm,
area using Eq. 1 (see Methods). (I) Recovered denoised topography map after quadric fitting (same area as in (g)). Color scale is same as for (g). (J) Localization precision of the Z
positions calculated as the error weighed standard error of the mean for a 3 × 3 pixel, 90 nm × 90 nm, area of the denoised topography maps. (K) Topography map from (i) overlaid with mean
membrane curvature. Source data EXTENDED DATA FIG. 4 VALIDATION OF RECOVERED MEMBRANE TOPOGRAPHY WITH RICM. (A) Illustration of the overlay of the reconstructed 3D topography map with the
corresponding reflection interference contrast microscopy (RICM) image. (B) Recovered membrane height plotted against RICM intensity for a representative cell. As predicted analytically by
theoretical models, membrane height and RICM intensity follow a co-sinusoidal relationship (see Methods). The correlation is fitted with a cosine function that describes the data with an R2
= 0.999. Data is binned using an error weighted rolling average (10 ± 10 nm) with error bars showing the s.e.m. Data is from N = 59,280 data points, _n_ = 20 cells from _n_ = 4 experiments.
Source data EXTENDED DATA FIG. 5 ASSESSMENT OF THE MEMBRANE STABILITY OVER TIME IMAGED WITH CONFOCAL. (A) Individual XZ images of a HEK293 acquired on the same location at t = 1 s, 20 s, 40
s, and 60 s. Initial visual assessment shows no major membrane displacements occur over t = 60 s. Data is from _n__R_ = 3. Scale bar, 1.5 μm. (B) Map of recovered Z position for the same
XZ-slice imaged over t = 60 s. (C) To assess membrane movement, a rolling standard deviation is calculated for each X position over a 6 s time window showing the average movement of the
membrane within the time frame needed for stable imaging. This allows us to image the spatial distribution of temporal nanoscopic displacements across the plasma membrane. (D) Histograms of
rolling standard deviation (grey) as calculated in (c) and accuracy of retrieving Z position of the membrane after quadric fitting (pink) (Extended Data Fig. 7e). (E) Median membrane
displacement, that is median of the rolling standard deviation, as a function of the applied time window. Pink dashed line represents the median accuracy in retrieving the Z position of the
membrane after quadric fitting. Error bars show s.d. Source data EXTENDED DATA FIG. 6 VALIDATION OF Β1AR DENSITY BY RATIO-METRIC IMAGING WITH A CELL MEMBRANE PROBE. (A) Schematic of
experimental approach. HEK293 cells express SNAP-β1AR and are labelled with SS488, while the membrane is labelled with CellMask DeepRed. (B) β1AR density normalized by recovered membrane
surface area versus β1AR density normalized by CellMask intensity. The dashed purple line is a linear fit to the data. Data is shown as error-weighted bins with equal number of data points
per bin. Error bars, s.e.m for y-axis and s.d. for x-axis. (C) Normalized β1AR density versus mean curvature. Recovery of normalized β1AR density by surface normalization (green) or by
CellMask intensity (purple) results in the same density-curvature correlation. Data is binned using an error-weighted rolling average (0.1 ± 0.1 μm−1) with error bars showing s.e.m. Data is
from _n_ = 7 cells from _n_ = 1 experiment. (D-E) 2D projection of topography-corrected and normalized density of CellMask and β1AR for the same region at the PM. CellMask density is uniform
at the PM (d), whereas β1AR forms domains (e). Scalebar, 500 nm. Source data EXTENDED DATA FIG. 7 NEITHER HIGH-DENSITY NOR LOW-DENSITY ACTIN ZONES ASSOCIATE WITH DOMAINS OF WELL-DEFINED
TOPOGRAPHY OR MEAN CURVATURE. (A) Actin intensity at the plasma membrane overlaid with boundaries of high- and low-density zones of actin. Scale bar, 500 nm. (B) Density map of normalized
β1AR overlaid with the actin boundaries from (a). Domain 1 in (a) and (b) indicates a low-actin-density region that contains both β1AR-enriched and -depleted domains. (C) Colocalization
analysis of high- and low-density zones of actin with β1AR-enriched and -depleted domains. The colocalization percentages are compared to those of randomized actin zones. Quantitative
correlations between high-/low-density actin regions and β1AR density patterns were either statistically nonsignificant or had low significance (_P_ = 0.06 n.s., _P_ = 0.03). _P_ values are
calculated by a two-sided paired _t_ test. Data are the mean ± s.d. for n_C_ = 23, n_R_ = 2. (D) Membrane topography overlaid with the actin boundaries from (a). High- and low actin density
zones are not preferentially colocalizing with membrane peaks or valleys. (E) Mean curvature overlaid with actin boundaries from (a). There is no preferential overlap of high- and low actin
zones with positive or negative curvatures. Overlays are representative for _n_ = 23 cells and _n_ = 2 experiments. Source data EXTENDED DATA FIG. 8 ACTIVATION BY AGONIST INDUCES CLATHRIN
REDISTRIBUTION FROM RECEPTOR-DEPLETED DOMAINS TO BOTH RECEPTOR-ENRICHED AND -DEPLETED DOMAINS. (A) Representative XY micrographs of SNAP-β1AR (magenta) and pmKate2-clathrin (green) and a
merge in HEK293 cells. (B-C) XZ micrograph of β1AR, clathrin and a merge. Clathrin colocalizes with depleted domains of β1AR (b), but not with β1AR-enriched domains in the apo state (c).
(D-F) XZ micrographs of events of β1AR internalization via clathrin-mediated endocytosis (indicated by arrows) after addition of ISO. These events are removed in our image analysis pipeline
for recovering membrane topography and GPCR density. For (b-f) β1AR is imaged in 3D STED (magenta), clathrin in confocal (green) and a merge is shown. (G-H) 2D projection of normalized
clathrin (g) and β1AR density (h) in the apo state. Arrows indicate regions of clathrin colocalizing with β1AR-depleted and not with β1AR-enriched domains. (I-J) 2D projection of normalized
clathrin (i) and β1AR density (j) after activation by ISO. Arrows indicate regions of clathrin colocalizing both with β1AR-enriched and -depleted domains, however β1AR-enriched and -depleted
domains do not always colocalize with clathrin (arrow with asterisk). (K) Colocalization analysis of high-density clathrin regions with β1AR-enriched and -depleted domains before and after
activation by isoproterenol (ISO). The colocalization percentages are compared to randomized clathrin zones. Under basal conditions, β1AR and clathrin were anticorrelated (_P_ = 0.01), while
after activation with the agonist ISO, clathrin colocalization with GPCR-enriched domains was not statistically significant (_P_ = 0.65). _P_ values are calculated by a two-sided paired _t_
test. Data are mean ± s.d. for n_C_ = 18, n_R_ = 3. Scalebar, (a) 1 µm, (b-f) 200 nm, (g-j) 500 nm. Source data EXTENDED DATA FIG. 9 CHANGES IN MEMBRANE TOPOGRAPHY, CURVATURE AND Β1AR
DENSITY AFTER CELL FLATTENING WITH AN AGAROSE PAD. Change in height (A), mean membrane curvature (B) and β1AR density (C) calculated by subtracting the values after flattening from the
values before flattening (Fig. 3c,e,g and d,f,h). (D) Quantification of absolute change in height after flattening with an agarose pad from data in (a). The red line indicates the average
change in membrane height of 14.5 nm. (E) Comparison of histograms of mean curvature of unperturbed (blue) and flattened (orange) for a single, representative HEK293 cell. After flattening
the width of the histogram is smaller compared to the unperturbed cells. (F) Histograms of normalized β1AR density before (blue) and after (orange) flattening of the same cell as in (e).
Normalized density of compressed cells shifts towards unity and the width of the histogram decreases. Data is representative for _n_ = 10 cells in _n_ = 5 replicates. EXTENDED DATA FIG. 10
DOMAINS OF H-RAS, PIEZO1 AND EGFR AT THE PM OF LIVE CELLS. 2D projection of H-Ras density (a), overlay of H-Ras density on super-resolved topography map (b) and mean curvature map (c) of
membrane topography shown in (b). H-Ras-enriched domains are formed at negative shallow curvature (see also Fig. 5d). (D-F) 2D projection of Piezo1 density (d), overlay of Piezo1 density on
super-resolved topography map (e) and mean curvature map (f). Piezo1-enriched domains have high contrast and strongly couple to positive shallow curvature (see Fig. 5e). (G-I) 2D projection
of EGFR density (g), overlay of EGFR density on super-resolved topography map (h) and mean curvature map (i). EGFR density variations couple to positive shallow curvature and are similar to
those of β1AR and other studied GPCRs (see Supplementary Fig. 23). Black arrows indicate examples of H-Ras-, Piezo1- and EGFR-enriched domains. Scalebar (a,d,g), 500 nm. SUPPLEMENTARY
INFORMATION SUPPLEMENTARY INFORMATION Supplementary Figs. 1–24 and Note. REPORTING SUMMARY SUPPLEMENTARY VIDEO 1 _xzy_ scanning for the reconstruction of superresolved membrane topography.
The movie shows the principle of reconstructing membrane topography. The _xz_ frames are scanned along the _y_ axis, and the blue line represents the recovered membrane height for each frame
SUPPLEMENTARY VIDEO 2 Overlay of recovered _z_ position on an _xzy_ stack imaged with confocal and 3D STED microscopy. The _xz_ frames are shown while scanning in the _y_ direction for
confocal (top) and 3D STED (bottom) imaging modes. The confocal _xz_ frames are overlaid with the recovered _z_ position of the membrane in confocal (yellow), whereas the 3D STED _xz_ frames
are overlaid with the recovered _z_ position in 3D STED (blue) and confocal (yellow). The recovered _z_ position accurately follows the variations of the PM for both confocal and 3D STED
imaging modes. SOURCE DATA SOURCE DATA FIG. 1 Statistical source data. SOURCE DATA FIG. 2 Statistical source data. SOURCE DATA FIG. 3 Statistical source data. SOURCE DATA FIG. 4 Statistical
source data. SOURCE DATA FIG. 5 Statistical source data. SOURCE DATA EXTENDED DATA FIG. 2 Statistical source data. SOURCE DATA EXTENDED DATA FIG. 3 Statistical source data. SOURCE DATA
EXTENDED DATA FIG. 4 Statistical source data. SOURCE DATA EXTENDED DATA FIG. 5 Statistical source data. SOURCE DATA EXTENDED DATA FIG. 6 Statistical source data. SOURCE DATA EXTENDED DATA
FIG. 7 Statistical source data. SOURCE DATA EXTENDED DATA FIG. 8 Statistical source data. RIGHTS AND PERMISSIONS Springer Nature or its licensor (e.g. a society or other partner) holds
exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely
governed by the terms of such publishing agreement and applicable law. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Kockelkoren, G., Lauritsen, L., Shuttle, C.G. _et al._
Molecular mechanism of GPCR spatial organization at the plasma membrane. _Nat Chem Biol_ 20, 142–150 (2024). https://doi.org/10.1038/s41589-023-01385-4 Download citation * Received: 13
January 2022 * Accepted: 14 June 2023 * Published: 17 July 2023 * Issue Date: February 2024 * DOI: https://doi.org/10.1038/s41589-023-01385-4 SHARE THIS ARTICLE Anyone you share the
following link with will be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer
Nature SharedIt content-sharing initiative




