- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT By performing whole-genome sequencing in a Swedish cohort of 71 parent-offspring trios, in which the child in each family is affected by systemic lupus erythematosus (SLE, OMIM
152700), we investigated the contribution of de novo variants to risk of SLE. We found de novo single nucleotide variants (SNVs) to be significantly enriched in gene promoters in SLE
patients compared with healthy controls at a level corresponding to 26 de novo promoter SNVs more in each patient than expected. We identified 12 de novo SNVs in promoter regions of genes
that have been previously implicated in SLE, or that have functions that could be of relevance to SLE. Furthermore, we detected three missense de novo SNVs, five de novo insertion-deletions,
and three de novo structural variants with potential to affect the expression of genes that are relevant for SLE. Based on enrichment analysis, disease-affecting de novo SNVs are expected
to occur in one-third of SLE patients. This study shows that de novo variants in promoters commonly contribute to the genetic risk of SLE. The fact that de novo SNVs in SLE were enriched to
promoter regions highlights the importance of using whole-genome sequencing for identification of de novo variants. SIMILAR CONTENT BEING VIEWED BY OTHERS HIGH-THROUGHPUT IDENTIFICATION OF
FUNCTIONAL REGULATORY SNPS IN SYSTEMIC LUPUS ERYTHEMATOSUS Article Open access 09 August 2024 TRANSMISSION DISEQUILIBRIUM ANALYSIS OF WHOLE GENOME DATA IN CHILDHOOD-ONSET SYSTEMIC LUPUS
ERYTHEMATOSUS Article 24 July 2023 THE _ZNF76_ RS10947540 POLYMORPHISM ASSOCIATED WITH SYSTEMIC LUPUS ERYTHEMATOSUS RISK IN CHINESE POPULATIONS Article Open access 04 March 2021 INTRODUCTION
Systemic lupus erythematosus (SLE, OMIM 152700) is a chronic autoimmune disease that affects multiple organs of the human body. SLE is ninefold enriched among women and has an estimated
prevalence in the Swedish population of 68 in 100,000 [1] and prevalence estimates vary between 20 and 100 per 100,000 in other European populations [2]. In the past decade, genome-wide
association studies have identified more than 100 risk loci that are associated with SLE [3, 4] and account for ~30% of the genetic susceptibility of SLE [5]. Mutations causing monogenic SLE
that contribute to the heritability of the disease have been detected in 1–3% of SLE patients [6]. In an earlier study we have shown that ultra-rare missense variants in the same genes as
those causing monogenic forms of SLE make an additional contribution to the heritability of SLE [7]. De novo variants (DNVs) could be an alternative mechanism that increases the burden of
genetic risk in SLE. De novo mutations occur during the formation of the gametes and are thereby not inherited from the parents, although they are present in every cell of the child. On
average 70 DNVs are expected to be introduced per human generation, and out of them one to two DNVs affect the protein-coding sequence [8]. Using whole-genome sequencing (WGS) of
parent-offspring trios both inherited mutations and DNVs can be detected. Most studies on DNVs in complex diseases have focused on developmental disorders like autism spectrum disorder and
schizophrenia [9], but also congenital heart defects have been found to be frequently caused by DNMs [10]. A majority of the sporadic cases that receive a molecular diagnosis by clinical
sequencing can be fully explained by de novo variants [11]. It has also been shown that the genomes of individuals with severe undiagnosed developmental disorders carry an excess of DNVs in
developmentally important genes. Based on exome sequencing it has been estimated that more than 40% of children with developmental diseases or syndromes carry deleterious DNVs in their
protein-coding sequences [12]. In addition, an enrichment of DNVs with incomplete penetrance has been observed in individuals with milder forms of autism [13]. DNVs have also been implicated
in certain sporadic cancers of young individuals [14]. As there is no selection pressure on non-lethal DNVs, the probability that the DNVs cause increased disease risk is higher than for
inherited variants in a population. The contribution of DNVs to genetic risk in complex diseases like SLE has not been investigated in depth, but as SLE is typically caused by a combination
of multiple genetic risk factors with small effect sizes, the disease contribution by SLE can be expected to be lower than for early-onset developmental disorders. The investigations of DNVs
in autoimmune diseases have so far focused on individuals with early-onset and severe cases of the disease [15], where the effects of de novo variants are presumed to be large and thus
easier to identify than in patients with the more common complex disease etiology. To identify DNVs in SLE, an early study used exome sequencing of 30 early-onset SLE patients with
extreme-phenotype, but this study only confirmed one de novo variant, in addition to 13 potential missense DNV in genes that are not expected to increase the risk of SLE [16]. An unbiased
and comprehensive investigation of the contribution of DNVs to SLE and other autoimmune disease still remains to be performed. RESULTS In this study the goal was to identify de novo single
nucleotide variants (SNVs), de novo insertion-deletions (INDELs) and de novo structural variants (SVs) in trio families with a child affected by SLE and to determine the potential
contribution of de novo variants to the genetic risk of SLE. We investigated the contribution of de novo variants to SLE using WGS of 71 parent-offspring trios. FREQUENCY AND ORIGIN OF DE
NOVO SINGLE NUCLEOTIDE VARIANTS IN SLE By using WGS to generate 30x sequencing coverage of the genomes of 213 individuals from 71 trio families, followed by calling of SNVs, stringent
filtering, and experimental validation of a subset of de novo SNVs by the polymerase chain reaction, followed by Sanger sequencing, we detected a total of 4157 de novo SNVs that were present
in the genomes of the child, but absent from the parents’ genomes. The total number of de novo SNVs detected in 71 SLE patients corresponds to on average 59 de novo SNVs per SLE patient
(range 40–91). This number compares well with the rate of 73 de novo SNVs detected per healthy individual in the Danish population using WGS combined with de novo assembly of the genomes of
25 healthy trio families [17], and with the data from a large study of 1548 trio families from the Icelandic population, which identified 65 de novo SNVs per trio [8]. Thus we do not find
evidence in our data that the total number of de novo SNVs per individual would be overrepresented in SLE compared with controls. The parent of origin of de novo SNVs in the offspring was
determined and, as expected, due to the continued cell divisions in sperm cells after puberty, the number of de novo SNVs originating from the father was almost four times higher (_n_ = 269)
than the number of de novo SNVs with a maternal origin (_n_ = 73). The effect of the age at conception of the father in our study was calculated to cause an increase of 1.56 de novo
variants per year in the child affected with SLE. This value corresponds to a correlation coefficient of 0.49 (_p_ value = 2.97E–7) between age of the father and the rate of de novo variants
(Supplementary Fig. S1). When conditioning the linear regression on the age of the mother, each year in the age of the father adds 1.34 de novo variants (_p_ value 0.00158). ENRICHMENT OF
DE NOVO SNVS TO PROMOTER REGIONS IN SLE PATIENTS Next we investigated whether de novo SNVs in SLE were enriched or depleted in specific genomic contexts. Comparison of the genomic
distribution of de novo SNVs in the SLE patients in our study with that of healthy individuals from an Icelandic study [8] allowed identification of regions of the genome that are
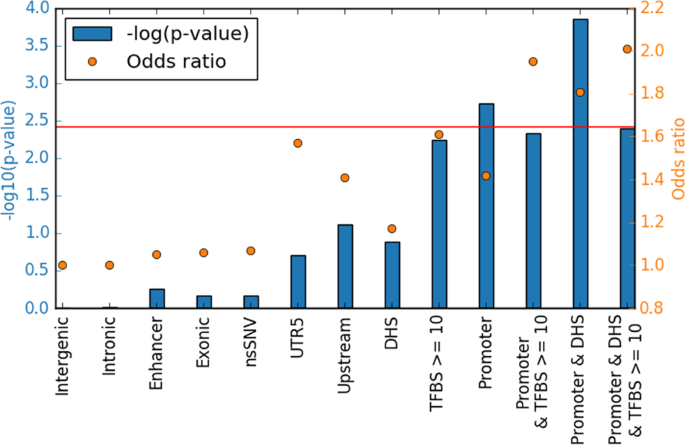
differentially affected by de novo SNVs in SLE. These regions could potentially be involved in the pathogenesis of SLE. For SNVs in functionally inactive regions, an odds ratio of 1.0 is
expected, which is in line with an odds ratio of 1.0 that we observe for the de novo SNVs in functionally inactive intronic and intergenic regions (Fig. 1). Compared with the de novo
variants in the data from the healthy individuals we observed a suggestive enrichment of de novo SNVs in 5′-untranslated regions (UTR5) and regions upstream of gene promoters (Table 1, Fig.
1), while de novo variants in SLE patients were not enriched in exons and enhancers. By annotation of the variants based on predicted chromatin states [18] in B lymphocytes, we observed a
significant enrichment of de novo SNVs in promoters in the SLE patients compared with healthy individuals (OR = 1.42, _p_ value 0.0019, Pearson’s chi-squared test). B cells are of particular
interest in SLE as the disease is characterized by B cell dysfunction [19]. The signal of enrichment was further strengthened by overlapping the DNV annotated to promoter regions with DHS
regions of open chromatin (1.81, _p_ value 0.00014). Small additional gains in odds ratio were also observed by adding a filter on variants that overlap more than 10 transcription factor
binding sites (TFBS) (Table 1, Fig. 1, and Supplementary Fig. S2). DE NOVO SNVS IN PROMOTERS WITH POTENTIAL TO AFFECT RISK OF SLE The genes located closest to the de novo SNVs identified in
promoter regions of the SLE genomes were investigated further regarding their putative roles as risk factors for SLE. Of the 87 de novo promoter SNVs only two were annotated to the same
genes (_ZNF292_). Pathway analysis of these genes using Gene Set Enrichment Analysis and the Genotype-Tissue Expression dataset revealed no significant pathways. Each of 33 of the 87 de novo
SNVs annotated to a promoter region were judged to have large potential to affect transcription factor binding due its their central position in relation to a large number of TFBS, its
closeness to a sequence peak of active histone marks and overlap with a region of open chromatin in B-lymphocytes [20]. Indeed, among these 33 SNVs, 25 were located close to genes with
differential gene expression between SLE patients and controls, according to data in the gene expression omnibus (GEO) database (FDR-corrected _p_ value < 0.01, Supplementary Table S1)
[21, 22], which compared with a random selection of de novo SNVs with matching properties and annotations in the healthy controls, this represents a significant enrichment (OR 1.66, _p_
value 0.0157). By literature search of the genes close to the 33 de novo SNVs located in central parts of a promoter we found twelve genes with known functions that could be relevant for SLE
directly or indirectly through involvement in autoimmunity, apoptosis, or inflammation (Table 2, Supplementary Table S2, Supplementary Fig. S3). None of the genes reported in Table 2 have
any similarly located de novo SNVs in the Icelandic controls. We also identified 9 additional de novo SNVs with high potential to affect the expression of a nearby gene, but where a
biological connection to SLE was not obvious (Supplementary Table S3, Supplementary Fig. S4). In addition to de novo SNVs in gene promoters, three nonsynonymous de novo SNVs located in
coding regions of the _MAZ, LTB4R2,_ and _ISX_ genes were identified. None of these genes had any nonsynonymous de novo SNVs in the controls. These nsSNVs affect zinc binding to the protein,
truncate the protein or affect the DNA-binding, respectively, and thereby most likely affect the function of these proteins (Table 3, Supplementary Table S4). _LTB4R2_ is a receptor for the
inflammation mediator leukotriene B4 that have been found to be greatly elevated in serum from SLE patients compared with healthy controls [23]. The other two genes play a role in the
immune system, but have not previously been implicated in the etiology of SLE. DE NOVO INSERTION-DELETIONS AND STRUCTURAL VARIANTS IN SLE PATIENTS De novo INDELs are more infrequent than
SNVs, but could have larger effects on gene regulation and protein function than SNVs, for example by introducing frameshifts. The low frequency of INDELs makes this study underpowered for
detection of INDEL enrichment in regions or genes that are relevant for SLE. Despite this limitation we detected 5 de novo INDELs that were located in the vicinity of the _TPR, CFLAR,
RACGAP1, ARID3B, GSDMD_ genes that are relevant for SLE (Supplementary Table S5). Structural variants have even greater potential than SNVs and INDELs to affect the function of proteins due
to their larger size. However, de novo SVs are infrequent in the human genome, with an expected frequency based on the data from the Icelandic population of ~13 large de novo SVs in our
entire trio dataset [24]. Initially, we identified 97 candidate de novo SVs in the SLE patients. Based on their genomic annotations and manual inspection of the sequence reads in the
integrated genome browser (IGV) 10 high confidence variants remained for further investigation. Of these 10 SVs, three SVs were close to genes or in genes that could contribute to increased
risk of SLE, see Table 4 and Supplementary Table S6. Two of these three are involved in IFN-kappaB signaling. DISCUSSION As DNMs are not inherited, they do not explain any of the “missing
heritability” in SLE, but they may contribute to the genetic risk of obtaining SLE in individual patients. We identified 12 de novo SNVs that may confer risk of SLE based on their position
in active gene promoters and based on literature searches of the functions of the genes carrying de novo SNVs. Several of the candidate genes with de novo SNVs do not have a previously known
connection to SLE, and thus they represent potential novel risk genes for SLE. However, as our strategy to identify risk genes was based on their known functions related to SLE, there
should be a number of additional de novo SNVs than those high-lighted here with potential to increase the genetic risk of SLE. The enrichment of de novo SNVs in gene promoters in SLE
patients compared with the data from the healthy individuals in the Icelandic population amounts to 26 de novo SNVs more than expected by chance, which indicates that de novo SNVs in
promoters are a common factor for increasing the genetic risk of SLE. The finding of de novo SNV enrichment in promoters would benefit from replication in another SLE or autoimmune patient
cohort. The 12 most plausible de novo SNVs located close genes that are relevant for SLE identified here are distributed across 11 patients. As there are no signs of clustering of de novo
SNVs in promoters within individual SLE patients, disease affecting de novo SNVs can be expected to occur in about one-third of SLE patients. In our study, we found 21 de novo SNV more than
expected by chance that are located in promoters overlapping DHS in the SLE patients compared with controls. Assuming a uniform probability distribution of the de novo SNVs between samples,
there are 18 SLE patients that have at least one potentially risk-contributing de novo SNV, which correspond to a frequency of 0.26 in our set of SLE patients and an odds ratio of 1.81
compared with controls. These numbers are comparable to for example the _IRF5_ locus which is one of the most strongly associated loci in SLE [3, 25]. This comparison suggests that de novo
SNVs may contribute significantly to the total burden of risk for SLE caused by genetic variation. The control data with de novo SNVs in the Islandic population was a valuable asset for our
study. The data are technically similar to our data in terms of sequencing coverage and methods used for read mapping and SNV calling. However, the Islandic project used three different
sequencing library preparation methods. In order to make the comparison of the de novo SNV data in healthy individuals from Iceland with our data from Swedish patients with SLE as precise as
possible, we filtered out de novo SNVs in overlapping regions with high GC-content in both data sets. We found that GC-content filtering had only a marginal effect on the enrichment and
significance of de novo SNVs, even in promoter regions which are GC-rich. Several SLE associated INDELs and SVs have previously been reported in SLE, which are exemplified by a 5 bp INDEL
close to _IRF5_ [26], copy number variation of _FCGR3A_ and _FCGR3B_ [27] in synergy with copy number variation of _ADAM3A_ [28], and copy number variation of _C4_ [29], as well as
population specific CNVs in _TLR7, DEFB4, RABGAP1L, and HLA-DRB5_ [28]. The CNVs that span protein-coding regions have a dose effect on the expressed proteins that correlates with the copy
number. The heterozygous deletion of several exons in _RBM10_ detected in this study will also have a dose effect as only one of the chromosomes will produce a functional protein. Rbm10
deficiency suppresses NF-κB-mediated responses in vivo in mouse models and in vitro [30], which supports role for the de novo SVs in increasing SLE risk as NF-κB-activity in T-cells from SLE
patients has been shown to be greatly decreased [31]. The fact that de novo SNVs are enriched to promoter regions and not in protein-coding exons highlights the importance of using WGS for
identification of de novo variants. However, in a larger patient cohort it is probable that also nonsynonymous de novo SNVs would be statistically significantly enriched. In this study, we
found only three potentially SLE risk contributing de novo SNVs that were located in protein coding regions, which is in line with a previous study that identified a small number of de novo
variants with large potential to affect the protein function by whole-exome sequencing of SLE trio families [16]. Although the missense de novo SNVs are uncommon they might confer a large
risk of SLE for the individual patient and therefore important for understanding the genetic risk underlying SLE. SLE is considered as a model for systemic autoimmune diseases because the
autoimmune process in SLE may affect most organs of the human body. Thus the finding of de novo variants in SLE could also be highly relevant for other systemic autoimmune diseases. METHODS
DNA SAMPLES FROM TRIO FAMILIES DNA was extracted from peripheral whole blood of 71 patients affected by SLE and their biological parents, who visited the rheumatology clinics in Uppsala (_n_
= 22), Karolinska Hospital (Stockholm) (_n_ = 30), Lund (_n_ = 15), and Linköping (_n_ = 4). All patients were examined by a rheumatologist and their medical records were reviewed. SLE
patients and their parents provided informed consent to participate in the study, and the study was approved by the regional ethics committees. Of the patients 85% were female, and on
average 24 years old (range 9–46) at onset of SLE. Fourteen of the patients were under 18 years old at SLE onset and could therefore be classified as having childhood-onset SLE. The patients
fulfilled at least four American College of Rheumatology (ACR) 1982 criteria for SLE [32], with the exception of five patients who displayed three ACR criteria together with a clinical
diagnosis of SLE. For further clinical data, see Supplementary Table S7. All patents included in the study were of European descent. WHOLE-GENOME SEQUENCING AND SEQUENCE ALIGNMENT Sequencing
libraries were prepared from 1 μg DNA using the TruSeq PCR-free DNA sample preparation kit (Illumina Inc.) targeting an insert size of 350 bp. 150 bp paired-end WGS was performed to >30
coverage using an Illumina HiSeqX sequencer with v2.5 chemistry. The sequences were aligned with BWA-MEM [33] version 0.7.12 using default parameters with the addition of the –M and –R flag,
and using the b37 human reference genome from the GATK file bundle version 2.8. The raw alignments where then flagged for duplication, realigned, and recalibrated using GATK version 3.3.0
[34]. Metrics from the WGS after mapping and variant calling are shown in Supplementary Table S8. CALLING OF SINGLE NUCLEOTIDE VARIANTS AND INSERTION-DELETIONS SNVs and INDELs in the WGS
data were called jointly for all samples using GATK version 3.5.0 following the GATK best practice protocol. In the variant recalibration step we used positive training data from the HapMap
project (with phred quality score prior likelihood of Q15, which is equal to 97% likelihood that the genotype is correct) and the 1000 Genomes Omni 2.5 M genotyping data (prior Q12, 94%
likelihood for correct genotypes) as well as in-house genotype data on the samples from the Infinium OmniExpressExome-8 v1.3 SNP chip (Illumina) with 958,497 SNP markers (prior Q20, 99%
likelihood). As additional training data we used the 1000 Genomes high confidence calls (prior Q10, 90% likelihood) and for annotation and statistics we used the dbSNP version 138 (prior Q2,
37% likelihood). All data files, except the in-house OmniExpressExome SNP genotype data, were obtained from the GATK file bundle version 2.8. Variants were marked as PASS if the variant
quality score log-odds were higher than that in the 99th percentile in the training data for both SNVs and INDELs. The variants were then further refined by calculating genotype posterior
likelihoods using the data from parent-offspring trios in GATK. Low quality variants were discarded if the genotype posterior had a score <Q20. CALLING AND FILTERING OF DE NOVO CANDIDATE
SINGLE NUCLEOTIDE VARIANTS AND INSERTION-DELETIONS De novo SNV and INDEL candidates were called using two programs: GATK version 3.5.0 and Triodenovo [35] version 0.04. By GATK de novo
variants were called with the VariantAnnotator using –annotation PossibleDeNovo and Triodenovo was run with default parameters. The program DNMFilter [36] v0.1.1 was used to discern true de
novo SNVs from false ones. DNMFilter assigns a score to each de novo SNV candidate that correlates with the probability that the candidate is a true de novo SNV. The score threshold was set
to 0.5. The model used by the program is based on the training data supplied in the program package using default features extracted from 264 trio samples. The raw de novo INDEL calls were
filtered and annotated as described in the Supplementary “Methods and Results”. VALIDATION OF DE NOVO SINGLE NUCLEOTIDE VARIANTS AND INSERTION-DELETIONS Primers for PCR were designed using
Primer 3 v4.0.0 [37], and the specificity of the primers was confirmed using the BLAST-like alignment tool [38] and in silico PCR. Whole-genome amplification of 50–100 ng of DNA was
performed using the REPLI-g Mini Kit (Qiagen), followed by specific PCRs using reagents from the Phusion Hot Start II kit. After amplification the PCR products with correct size were cleaned
using Exo/SAP reagents (GE healthcare), and the concentration and size of the PCR products were determined using Qubit HS (Invitrogen). Twenty ng of each PCR product were mixed with 4 pmol
of primer and subjected to Sanger sequencing on a Genetic Analyser (Life Technologies). The sequencing results for the SNVs were read using the program Sequence Scanner v2.0, whereas the
INDELs were analysed using Chromas Lite v2.3. To assess the performance of the de novo variant calling pipeline, 110 SNVs and 83 INDELs were randomly selected for validation by Sanger
sequencing in all family members. For the SNVs, the primer construction and the sequencing was successful for 99 of the primer pairs, out of which 98 were confirmed to be de novo variants,
which corresponds to an FDR of 1%. Sequencing of the INDELs returned results for 71 of the primer pairs, out of which 56 were confirmed to be de novo, which corresponds to an FDR of 21%. The
high accuracy of our data and the pipeline for calling de novo SNVs is demonstrated by an allele balance between 30 and 70% for 98% of the called the de novo SNVs after filtering [17, 39].
Thus the called de novo SNVs contained very few mosaic variants and few variants were called due to sequencing errors. Notably, before filtering the majority of the variant alleles were
outside of the 30–70% range, see Supplementary Fig. S5. In addition to the randomly selected variants, eight of the variants that could potentially confer risk for SLE (Table 2) were
confirmed to be de novo and none of them failed validation. The single potential disease related INDEL tested was also confirmed to be a de novo variant. CALLING AND FILTERING DE NOVO
STRUCTURAL VARIANTS Germline SVs in the form of INDELs larger than 10 base pairs were called by Manta-1.0.3 [40] in samples from parents and SLE patients using default parameters and
additionally by FermiKit-0.13 [41] in the patients using the parameter –s3g for genome size and 150 for read-length. Because Manta and FermiKit do not inherently support calling of de novo
variants we developed our own calling and filtering procedure for de novo SVs based on germline variant calls, with a focus on high specificity for variant calling. To call de novo variants
we first selected variants exceeding 10 base pairs in the SLE patients that were detected by both FermiKit and Manta using a reciprocal overlap of 50%. FermiKit and Manta complement each
other as they use different approaches to call SVs. Using only overlapping calls should therefore greatly reduce the number of false positives. Structural variants of the same type that
remained undetected, using a reciprocal overlap of 50%, in any of the trio family samples were kept. The overlap was calculated using both the actual called SVs and the candidate variants
generated by the programs. The remaining SVs were annotated similarly as for INDEL filtering, using annotations from blacklisted regions by ENCODE, mappability scores, segmental
duplications, and RepeatMasker. In addition, structural variant regions were removed if the coverage in the region flanking the variant followed a similar pattern between an SLE patient and
any of the two parents. The presence of the de novo SVs was also confirmed manually in the integrated genome viewer (IGV). FUNCTIONAL ANNOTATION OF DE NOVO SINGLE NUCLEOTIDE VARIANTS AND
INSERTION-DELETIONS The variants were annotated functionally using Annovar version 2016.05.11 [42] with the annotations data described below supplied to Annovar via the. Chromatin states
were annotated according to ChromHMM [18] predictions based on the B-lymphocyte cell line GM12878. Relative gene positions were obtained using RefSeq [43]. TFBS were annotated using the
chromatin-immunoprecipitaion sequencing (ChIP-seq)-based UCSC genome browser track wgEncodeRegTfbsClustered from ENCODE. DNase I hypersensitive site (DHS) annotations were based on the UCSC
genome browser track wgEncodeOpenChromDnaseGm12878Pk based on the GM12878 cell line from ENCODE. The effects of de novo nsSNVs on the functions of the encoded proteins were determined using
positional annotations from the UniProtKB [44], 3-dimensional protein structures in the protein data bank (PDB) [45] or structural models in the Protein Model Portal [46]. Gene expression
data of nearby genes was obtained from GeneCards, collected from the Human Integrated Protein Expression Database (www.genecards.org, [47, 48]). We also used the USCS genome browser to
manually inspected de novo SNVs in promoter regions in to find de novo SNVs with large potential to affect transcription factor binding due to their positions in relation to TFBS, histone
marks, and open chromatin in B-lymphocytes [20]. PARENT OF ORIGIN AND AGE AT CONCEPTION The parent of origin of de novo SNVs in the offspring was determined in the genomes of 32 trio family
members for those variants that had an additional informative SNV within the same sequencing read (_n_ = 342 de novo SNVs). At least two reads spanning both the additional SNV and the de
novo SNV supporting the same parent was required to assign the parental origin of a de novo SNV. The number of expected de novo SNVs in the offspring is affected by the age at conception of
the father and to a lesser extent the age of the mother [8]. The correlation and _p_ value of the correlation between age of the father and the child affected with SLE was calculated using
linear regression in R. DE NOVO SINGLE NUCLEOTIDE VARIANTS IN HEALTHY CONTROLS A set of 97,942 de novo SNV and over 30 million SNVs found in 1548 trios from a study in Iceland by deCODE
Genetics was downloaded from the European Variation Archive (www.ebi.ac.uk/eva/), study accession number PRJEB15197, to be used as healthy control data in our study [8]. The SNVs in the
Icelandic data were then annotated exactly as the de novo SNVs detected in the Swedish SLE trios. For the enrichment analysis we also filtered out de novo SNVs in overlapping regions with
high GC-content in both datasets. The SNVs filtered were located in 100 bp regions with at least 75% GC content or located in any of the 1000 “difficult promoters” defined by Ross et al.
[49]. Among the Icelandic de novo SNVs, 87 were randomly selected to match the number of SNVs found in the Swedish SLE patients. The de novo SNVs in Iceland were visually inspected in the
UCSC genome browser in an identical fashion as the de novo SNVs in the SLE patients to determine a subset of de novo SNVs most likely to affect the promoter. Enrichment _p_ values were
calculated using the Pearson’s chi-squared test with Yates’ correction. CALCULATION OF ODDS RATIOS AND SIGNIFICANCE IN SELECTED ANNOTATED REGIONS Odds ratios between de novo SNVs found in
our SLE cohort and the controls were calculated for selected chromatin states determined by ChIP-seq annotations in gene flanking regions and intergenic regions, DHS, number of TFBS, and
combinations of these regions. The significance of the enrichment was then calculated using Pearson’s chi-squared test. DIFFERENTIAL GENE EXPRESSION DATA SETS We used two publicly available
gene expression datasets deposited in GEO [50] to calculate FDR corrected _p_ values for differential expression between SLE patients and controls using the online service GEO2R
(https://www.ncbi.nlm.nih.gov/geo/geo2r/). One of the studies (GEO accession number: GSE45291) contains expression array data for 54715 probes from 292 SLE patients and 20 controls [21]. The
other study (GEO accession number: GSE65391) is a longitudinal study of pediatric SLE patients where the sample taken at the first visit to the clinic was used to calculate differential
gene expression for 43,798 probes between 134 SLE patients and 36 controls [22]. The lowest probe _p_ value for each gene from the two studies was used in our study. DATA AVAILABILITY De
novo SNVs and INDELs presented in this paper have been submitted to dbSNP (https://www.ncbi.nlm.nih.gov/projects/SNP/) under the submitter ID MOLMED_UU and Submitter batch ID
Denovo_variants_1.0 with the submission SNP IDs of ss2137544038 and ss3798736965 to ss3798737055. REFERENCES * Stahl-Hallengren C, Jonsen A, Nived O, Sturfelt G. Incidence studies of
systemic lupus erythematosus in Southern Sweden: increasing age, decreasing frequency of renal manifestations and good prognosis. J Rheumatol. 2000;27:685–91. CAS PubMed Google Scholar *
Rees F, Doherty M, Grainge MJ, Lanyon P, Zhang W. The worldwide incidence and prevalence of systemic lupus erythematosus: a systematic review of epidemiological studies. Rheumatol (Oxf).
2017;56:1945–61. Article Google Scholar * Langefeld CD, Ainsworth HC, Cunninghame Graham DS, Kelly JA, Comeau ME, Marion MC, et al. Transancestral mapping and genetic load in systemic
lupus erythematosus. Nat Commun. 2017;8:16021. Article CAS PubMed PubMed Central Google Scholar * Chen L, Morris DL, Vyse TJ. Genetic advances in systemic lupus erythematosus: an
update. Curr Opin Rheumatol. 2017;29:423–33. Article PubMed Google Scholar * Morris DL, Sheng Y, Zhang Y, Wang YF, Zhu Z, Tombleson P, et al. Genome-wide association meta-analysis in
Chinese and European individuals identifies ten new loci associated with systemic lupus erythematosus. Nat Genet. 2016;48:940–6. Article CAS PubMed PubMed Central Google Scholar *
Costa-Reis P, Sullivan KE. Monogenic lupus: it’s all new! Curr Opin Immunol. 2017;49:87–95. Article CAS PubMed Google Scholar * Almlof JC, Nystedt S, Leonard D, Eloranta ML, Grosso G,
Sjowall C, et al. Whole-genome sequencing identifies complex contributions to genetic risk by variants in genes causing monogenic systemic lupus erythematosus. Hum Genet. 2019;138:141–50.
Article PubMed PubMed Central CAS Google Scholar * Jonsson H, Sulem P, Kehr B, Kristmundsdottir S, Zink F, Hjartarson E, et al. Parental influence on human germline de novo mutations in
1,548 trios from Iceland. Nature. 2017;549:519–22. Article PubMed CAS Google Scholar * Veltman JA, Brunner HG. De novo mutations in human genetic disease. Nat Rev Genet. 2012;13:565–75.
Article CAS PubMed Google Scholar * Zaidi S, Choi M, Wakimoto H, Ma L, Jiang J, Overton JD, et al. De novo mutations in histone-modifying genes in congenital heart disease. Nature.
2013;498:220–3. Article CAS PubMed PubMed Central Google Scholar * Yang Y, Muzny DM, Xia F, Niu Z, Person R, Ding Y, et al. Molecular findings among patients referred for clinical
whole-exome sequencing. JAMA. 2014;312:1870–9. Article CAS PubMed PubMed Central Google Scholar * Deciphering Developmental Disorders S. Prevalence and architecture of de novo mutations
in developmental disorders. Nature. 2017;542:433–8. Article CAS Google Scholar * Neale BM, Kou Y, Liu L, Ma’ayan A, Samocha KE, Sabo A, et al. Patterns and rates of exonic de novo
mutations in autism spectrum disorders. Nature. 2012;485:242–5. Article CAS PubMed PubMed Central Google Scholar * Gonzalez KD, Buzin CH, Noltner KA, Gu D, Li W, Malkin D, et al. High
frequency of de novo mutations in Li-Fraumeni syndrome. J Med Genet. 2009;46:689–93. Article CAS PubMed Google Scholar * Duncan CJA, Dinnigan E, Theobald R, Grainger A, Skelton AJ,
Hussain R, et al. Early-onset autoimmune disease due to a heterozygous loss-of-function mutation in TNFAIP3 (A20). Ann Rheum Dis. 2018;77:783–6. Article CAS PubMed Google Scholar *
Pullabhatla V, Roberts AL, Lewis MJ, Mauro D, Morris DL, Odhams CA, et al. De novo mutations implicate novel genes in systemic lupus erythematosus. Hum Mol Genet. 2018;27:421–9. Article CAS
PubMed Google Scholar * Besenbacher S, Liu S, Izarzugaza JM, Grove J, Belling K, Bork-Jensen J, et al. Novel variation and de novo mutation rates in population-wide de novo assembled
Danish trios. Nat Commun. 2015;6:5969. Article CAS PubMed Google Scholar * Ernst J, Kellis M. ChromHMM: automating chromatin-state discovery and characterization. Nat Methods.
2012;9:215–6. Article CAS PubMed PubMed Central Google Scholar * Karrar S, Cunninghame Graham DS. Abnormal B cell development in systemic lupus erythematosus: what the genetics tell us.
Arthritis Rheumatol. 2018;70:496–507. Article PubMed PubMed Central Google Scholar * Kent WJ, Sugnet CW, Furey TS, Roskin KM, Pringle TH, Zahler AM, et al. The human genome browser at
UCSC. Genome Res. 2002;12:996–1006. Article CAS PubMed PubMed Central Google Scholar * Petri M, Fu W, Ranger A, Allaire N, Cullen P, Magder LS, et al. Association between changes in
gene signatures expression and disease activity among patients with systemic lupus erythematosus. BMC Med Genomics. 2019;12:4. Article PubMed PubMed Central Google Scholar * Banchereau
R, Hong S, Cantarel B, Baldwin N, Baisch J, Edens M, et al. Personalized immunomonitoring uncovers molecular networks that stratify lupus patients. Cell. 2016;165:551–65. Article CAS
PubMed PubMed Central Google Scholar * Wu T, Xie C, Han J, Ye Y, Weiel J, Li Q, et al. Metabolic disturbances associated with systemic lupus erythematosus. PLoS One. 2012;7:e37210.
Article CAS PubMed PubMed Central Google Scholar * Brandler WM, Antaki D, Gujral M, Noor A, Rosanio G, Chapman TR, et al. Frequency and complexity of de novo structural mutation in
autism. Am J Hum Genet. 2016;98:667–79. Article CAS PubMed PubMed Central Google Scholar * Sigurdsson S, Nordmark G, Goring HH, Lindroos K, Wiman AC, Sturfelt G, et al. Polymorphisms in
the tyrosine kinase 2 and interferon regulatory factor 5 genes are associated with systemic lupus erythematosus. Am J Hum Genet. 2005;76:528–37. Article CAS PubMed PubMed Central Google
Scholar * Sigurdsson S, Goring HH, Kristjansdottir G, Milani L, Nordmark G, Sandling JK, et al. Comprehensive evaluation of the genetic variants of interferon regulatory factor 5 (IRF5)
reveals a novel 5 bp length polymorphism as strong risk factor for systemic lupus erythematosus. Hum Mol Genet. 2008;17:872–81. Article CAS PubMed Google Scholar * Qi Y, Zhou X, Bu D,
Hou P, Lv J, Zhang H. Low copy numbers of FCGR3A and FCGR3B associated with Chinese patients with SLE and AASV. Lupus. 2017;26:1383–9. Article CAS PubMed Google Scholar * Barbosa FB,
Simioni M, Wiezel CEV, Torres FR, Molck MC, Bonilla MM, et al. Copy number variation in the susceptibility to systemic lupus erythematosus. PLoS ONE. 2018;13:e0206683. Article PubMed
PubMed Central CAS Google Scholar * Chen JY, Wu YL, Mok MY, Wu YJ, Lintner KE, Wang CM, et al. Effects of complement C4 gene copy number variations, size dichotomy, and C4A deficiency on
genetic risk and clinical presentation of systemic lupus erythematosus in East asian populations. Arthritis Rheumatol. 2016;68:1442–53. Article CAS PubMed PubMed Central Google Scholar
* Atsumi T, Suzuki H, Jiang JJ, Okuyama Y, Nakagawa I, Ota M, et al. Rbm10 regulates inflammation development via alternative splicing of Dnmt3b. Int Immunol. 2017;29:581–91. Article CAS
PubMed Google Scholar * Wong HK, Kammer GM, Dennis G, Tsokos GC. Abnormal NF-kappa B activity in T lymphocytes from patients with systemic lupus erythematosus is associated with decreased
p65-RelA protein expression. J Immunol. 1999;163:1682–9. CAS PubMed Google Scholar * Tan EM, Cohen AS, Fries JF, Masi AT, McShane DJ, Rothfield NF, et al. The 1982 revised criteria for
the classification of systemic lupus erythematosus. Arthritis Rheum. 1982;25:1271–7. Article CAS PubMed Google Scholar * Li H, Durbin R. Fast and accurate short read alignment with
Burrows-Wheeler transform. Bioinformatics. 2009;25:1754–60. Article CAS PubMed PubMed Central Google Scholar * McKenna A, Hanna M, Banks E, Sivachenko A, Cibulskis K, Kernytsky A, et
al. The Genome Analysis Toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010;20:1297–303. Article CAS PubMed PubMed Central Google Scholar
* Wei Q, Zhan X, Zhong X, Liu Y, Han Y, Chen W, et al. A Bayesian framework for de novo mutation calling in parents-offspring trios. Bioinformatics. 2015;31:1375–81. Article CAS PubMed
Google Scholar * Liu Y, Li B, Tan R, Zhu X, Wang Y. A gradient-boosting approach for filtering de novo mutations in parent-offspring trios. Bioinformatics. 2014;30:1830–6. Article CAS
PubMed PubMed Central Google Scholar * Untergasser A, Cutcutache I, Koressaar T, Ye J, Faircloth BC, Remm M, et al. Primer3–new capabilities and interfaces. Nucleic Acids Res.
2012;40:e115. Article CAS PubMed PubMed Central Google Scholar * Kent WJ. BLAT–the BLAST-like alignment tool. Genome Res. 2002;12:656–64. Article CAS PubMed PubMed Central Google
Scholar * Kosmicki JA, Samocha KE, Howrigan DP, Sanders SJ, Slowikowski K, Lek M, et al. Refining the role of de novo protein-truncating variants in neurodevelopmental disorders by using
population reference samples. Nat Genet. 2017;49:504–10. Article CAS PubMed PubMed Central Google Scholar * Chen X, Schulz-Trieglaff O, Shaw R, Barnes B, Schlesinger F, Kallberg M, et
al. Manta: rapid detection of structural variants and indels for germline and cancer sequencing applications. Bioinformatics. 2016;32:1220–2. Article CAS PubMed Google Scholar * Li H.
FermiKit: assembly-based variant calling for Illumina resequencing data. Bioinformatics. 2015;31:3694–6. CAS PubMed PubMed Central Google Scholar * Wang K, Li M, Hakonarson H. ANNOVAR:
functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 2010;38:e164. Article PubMed PubMed Central CAS Google Scholar * Pruitt KD, Tatusova
T, Maglott DR. NCBI reference sequences (RefSeq): a curated non-redundant sequence database of genomes, transcripts and proteins. Nucleic Acids Res. 2007;35:D61–5. Article CAS PubMed
Google Scholar * The UniProt C: UniProt. the universal protein knowledgebase. Nucleic Acids Res. 2017;45:D158–69. Article CAS Google Scholar * Berman HM, Westbrook J, Feng Z, Gilliland
G, Bhat TN, Weissig H, et al. The protein data bank. Nucleic Acids Res. 2000;28:235–42. Article CAS PubMed PubMed Central Google Scholar * Haas J, Roth S, Arnold K, Kiefer F, Schmidt T,
Bordoli L, et al. The Protein Model Portal–a comprehensive resource for protein structure and model information. J Biol Databases Curation 2013;2013:bat031. Google Scholar * Stelzer G,
Rosen N, Plaschkes I, Zimmerman S, Twik M, Fishilevich S, et al. The GeneCards suite: from gene data mining to disease genome sequence analyses. Curr Protoc Bioinforma. 2016;54:1 30 31–31 30
33. Article Google Scholar * Fishilevich S, Zimmerman S, Kohn A, Iny Stein T, Olender T, Kolker E, et al. Genic insights from integrated human proteomics in GeneCards. Database.
2016;2016:baw030. * Ross MG, Russ C, Costello M, Hollinger A, Lennon NJ, Hegarty R, et al. Characterizing and measuring bias in sequence data. Genome Biol. 2013;14:R51. Article PubMed
PubMed Central Google Scholar * Barrett T, Wilhite SE, Ledoux P, Evangelista C, Kim IF, Tomashevsky M, et al. NCBI GEO: archive for functional genomics data sets–update. Nucleic Acids Res.
2013;41:D991–5. Article CAS PubMed Google Scholar Download references ACKNOWLEDGEMENTS Sequencing was performed by the SNP&SEQ Technology Platform in Uppsala. The facility is part
of the National Genomics Infrastructure (NGI) Sweden and Science for Life Laboratory. The SNP&SEQ Platform is also supported by the Swedish Research Council and the Knut and Alice
Wallenberg Foundation. Sanger sequencing was performed by the Uppsala Genome Center Sequencing Facility. FUNDING This work was supported by grants from the Knut and Alice Wallenberg
Foundation, the Swedish Research Council for Medicine and Health 2018-02399 to LR and 2017-02000 to ACS, the Swedish Rheumatism Association, King Gustaf V’s 80-year Foundation and the
Swedish Society of Medicine and the Ingegerd Johansson donation. AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * Department of Medical Sciences, Molecular Medicine and Science for Life
Laboratory, Uppsala University, 751 23, Uppsala, Sweden Jonas Carlsson Almlöf, Sara Nystedt, Aikaterini Mechtidou & Ann-Christine Syvänen * Department of Medical Sciences, Rheumatology
and Science for Life Laboratory, Uppsala University, 751 85, Uppsala, Sweden Dag Leonard, Maija-Leena Eloranta, Lars Rönnblom & Johanna K. Sandling * Department of Medicine, Karolinska
Institutet, Rheumatology, Karolinska University Hospital, 171 77, Stockholm, Sweden Giorgia Grosso, Iva Gunnarsson & Elisabet Svenungsson * Department of Clinical and Experimental
Medicine, Rheumatology/Division of Neuro and Inflammation Sciences, Linköping University, 581 83, Linköping, Sweden Christopher Sjöwall * Department of Clinical Sciences, Rheumatology, Lund
University, Skåne University Hospital, 222 42, Lund, Sweden Anders A. Bengtsson & Andreas Jönsen Authors * Jonas Carlsson Almlöf View author publications You can also search for this
author inPubMed Google Scholar * Sara Nystedt View author publications You can also search for this author inPubMed Google Scholar * Aikaterini Mechtidou View author publications You can
also search for this author inPubMed Google Scholar * Dag Leonard View author publications You can also search for this author inPubMed Google Scholar * Maija-Leena Eloranta View author
publications You can also search for this author inPubMed Google Scholar * Giorgia Grosso View author publications You can also search for this author inPubMed Google Scholar * Christopher
Sjöwall View author publications You can also search for this author inPubMed Google Scholar * Anders A. Bengtsson View author publications You can also search for this author inPubMed
Google Scholar * Andreas Jönsen View author publications You can also search for this author inPubMed Google Scholar * Iva Gunnarsson View author publications You can also search for this
author inPubMed Google Scholar * Elisabet Svenungsson View author publications You can also search for this author inPubMed Google Scholar * Lars Rönnblom View author publications You can
also search for this author inPubMed Google Scholar * Johanna K. Sandling View author publications You can also search for this author inPubMed Google Scholar * Ann-Christine Syvänen View
author publications You can also search for this author inPubMed Google Scholar CORRESPONDING AUTHOR Correspondence to Jonas Carlsson Almlöf. ETHICS DECLARATIONS CONFLICT OF INTEREST The
authors declare that they have no conflict of interest. ADDITIONAL INFORMATION PUBLISHER’S NOTE Springer Nature remains neutral with regard to jurisdictional claims in published maps and
institutional affiliations. SUPPLEMENTARY INFORMATION SUPPLEMENTAL FIGURES S1,S2,S5 AND TABLES S1,S3,S5,S8 SUPPLEMENTAL FIGURE S3 SUPPLEMENTAL FIGURE S4 SUPPLEMENTAL TABLE S2 SUPPLEMENTAL
TABLE S4 SUPPLEMENTAL TABLE S6 SUPPLEMENTAL TABLE S7 SUPPLEMENTAL MATERIAL AND RESULTS RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative Commons Attribution 4.0
International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the
source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative
Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by
statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit
http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Almlöf, J.C., Nystedt, S., Mechtidou, A. _et al._ Contributions of de novo variants
to systemic lupus erythematosus. _Eur J Hum Genet_ 29, 184–193 (2021). https://doi.org/10.1038/s41431-020-0698-5 Download citation * Received: 16 January 2020 * Revised: 04 June 2020 *
Accepted: 14 July 2020 * Published: 28 July 2020 * Issue Date: January 2021 * DOI: https://doi.org/10.1038/s41431-020-0698-5 SHARE THIS ARTICLE Anyone you share the following link with will
be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt
content-sharing initiative







