- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT Glioblastoma (GBM) is a highly aggressive tumor with poor prognosis. A small subpopulation of glioma stem cells (GSCs) has been implicated in radiation resistance and tumor
recurrence. In this study we analyzed the expression of miRNAs associated with the functions of GSCs using miRNA microarray analysis of these cells compared with human neural stem cells.
These analyses identified gene clusters associated with glioma cell invasiveness, axonal guidance, and TGF-β signaling. miR-504 was significantly downregulated in GSCs compared with NSCs,
its expression was lower in GBM compared with normal brain specimens and further decreased in the mesenchymal glioma subtype. Overexpression of miR-504 in GSCs inhibited their self-renewal,
migration and the expression of mesenchymal markers. The inhibitory effect of miR-504 was mediated by targeting Grb10 expression which acts as an oncogene in GSCs and GBM. Overexpression of
exogenous miR-504 resulted also in its delivery to cocultured microglia by GSC-secreted extracellular vesicles (EVs) and in the abrogation of the GSC-induced polarization of microglia to M2
subtype. Finally, miR-504 overexpression prolonged the survival of mice harboring GSC-derived xenografts and decreased tumor growth. In summary, we identified miRNAs and potential target
networks that play a role in the stemness and mesenchymal transition of GSCs and the miR-504/Grb10 pathway as an important regulator of this process. Overexpression of miR-504 exerted
antitumor effects in GSCs as well as bystander effects on the polarization of microglia via delivery by EVs. SIMILAR CONTENT BEING VIEWED BY OTHERS MIR-146A-5P DEFICIENCY IN EXTRACELLULAR
VESICLES OF GLIOMA-ASSOCIATED MACROPHAGES PROMOTES EPITHELIAL-MESENCHYMAL TRANSITION THROUGH THE NF-ΚB SIGNALING PATHWAY Article Open access 30 June 2023 MICRORNA-27A-3P DELIVERED BY
EXTRACELLULAR VESICLES FROM GLIOBLASTOMA CELLS INDUCES M2 MACROPHAGE POLARIZATION VIA THE _EZH1/KDM3A/CTGF_ AXIS Article Open access 14 May 2022 INHIBITION OF HYPOXIC EXOSOMAL MIR-423-3P
DECREASES GLIOMA PROGRESSION BY RESTRICTING AUTOPHAGY IN ASTROCYTES Article Open access 08 April 2025 INTRODUCTION Glioblastomas (GBMs) are the most common and aggressive of the astrocytic
tumors. They are characterized by increased proliferation and angiogenesis, invasion into the surrounding normal tissue and resistance to therapies1. The prognosis of patients with GBM
remains extremely poor, and the median survival of GBM patients has remained around 14–16 months over the past decades2. Gene expression profiling studies identified GBM subtypes that are
classified based on their transcriptional signatures into various molecular groups, including proneural, classical, and mesenchymal3,4. Recently, the profiling of DNA methylation patterns in
glioma has refined these categories5, aligning them with the WHO 2016 diagnostic schema for glioma. These subtypes have distinct differential genetic alterations, molecular signatures,
cellular phenotypes, and patient prognosis5,6,7. GBMs contain a small subpopulation of cancer stem cells (i.e., glioma stem cells [GSCs])8 that are characterized by self-renewal,
multi-lineage differentiation potential, and the ability to generate xenografts that recapitulate the parental tumors9. GSCs have been implicated in tumor infiltration and resistance to
radio- and chemotherapy as well as tumor recurrence10. GSCs share stemness characteristics with neural stem cells (NSCs) but differ in their differentiation and oncogenic potentials9,11,12.
The epithelial-to-mesenchymal transition (EMT) is a process that allows epithelial cells to abandon their polarity and cell-to-cell adhesion properties and acquire mesenchymal cell
phenotypes which are associated with enhanced invasiveness, stemness and metastasis, and treatment resistance13. Recent studies demonstrated a similar process in glioma, and
proneural-to-mesenchymal transition in these tumors is characterized by increased aggressiveness, invasiveness and therapy resistance14,15,16. Mesenchymal transition can occur in recurrent
tumors and in response to radiation17,18 and is associated with poor patient prognosis. Tumor aggressiveness and mesenchymal transition of glioma are induced by cells and secreted factors in
the tumor microenvironment19,20,21. These cells include endogenous central nervous system (CNS) cells such as astrocytes, oligodendrocytes, neurons and microglia, and infiltrating immune
cells21. Microglia are resident immune cells in the brain and together with infiltrating macrophages represent the most prevalent CNS-associated cells in the tumors22. Microglia and
recruited macrophages have been reported to undergo differentiation to cells with M2-type characteristics, in response to factors secreted by glioma cells23. The M2 microglia and macrophages
further support tumor growth via the secretion of growth factors, chemokines and extracellular matrix-modifying enzymes22,24. In addition, recent studies implicated extracellular vesicles
(EVs) as important mediators of intercellular communication and in the cross talk of tumor cells and their microenvironments25. EVs contain proteins, lipids, and various RNA species and play
important roles in the interaction of glioma cells and microglia23,25. The expression of specific microRNAs (miRNAs) has been shown to be associated with several aspects of glioma
pathogenesis including cell cycle control, invasion, migration, resistance to therapies, and cell apoptosis26. Specific miRNAs have been also implicated in the self-renewal, stemness, and
tumorigenic features of GSCs27. Here, we analyzed the miRNA expression of GSCs in comparison with human NSCs (hNSCs) and identified unique miRNA expression profiles that distinguish these
two cell populations. Focusing on miR-504, which was highly expressed in hNSCs compared to GSCs, we demonstrated that it was upregulated in the G-CIMP glioma subtype compared to other GBM
subtypes. Moreover, we found that miR-504 exerted an antitumor effect in vitro and in vivo and in addition, inhibited the stemness and mesenchymal transit of GSCs. In addition,
overexpression of miR-504 in GSCs exerted a bystander effect on cocultured microglia cells by promoting the differentiation of these cells toward M1 phenotype via EV delivery. MATERIALS AND
METHODS GSC CULTURES All human materials were used in accordance with the policies of the Henry Ford Hospital Institutional Review Board. Generation of GSCs from fresh GBM specimens and
their characterization have been recently described28,29,30,31. Briefly, the GSCs were maintained in neurosphere medium (DMEM-F12 1:1, glutamine 10 mM, HEPES buffer 10 mM, and sodium
bicarbonate 0.025%) supplemented with basic fibroblast growth factor (20 ng/ml) and epidermal growth factor (20 ng/ml). The GSCs expressed markers such as CD44, CD133, Musashi-1, Sox2, and
nestin, exhibited self-renewal, and expressed astrocytic and neuronal markers upon differentiation. The GSCs also exhibited tumorigenic potential and generated glioma xenografts in nude
mice28,29,30,31,32,33. The full information of the GSCs employed in this study is described in Table S1. MICROGLIA AND NSC CULTURES Human microglial cells were obtained from Applied
Biological Material (Richmond, BC, Canada). All cells employed in this study were tested for mycoplasma contamination (Mycoplasma PCR Detection Kit) and found negative. hNSCs (H9, hESC
derived) were obtained from Invitrogen. TRANSDUCTION OF GSCS AND MICROGLIAL CELLS Lentivirus vectors (System Biosciences, Mountain View, CA) expressing the miR-504 reporter, pre-miR-504,
miR-504 antagomiR, Grb10, or control and Grb10 shRNAs were packaged and used to transduce the cells according to the manufacturer’s protocol and as previously described28,29,30. NEUROSPHERE
FORMATION ASSAY To determine the ability of GSCs to form secondary neurospheres, cells were plated in 24-well plates at a density of 10 and 100 cells/well through limiting dilution and the
number of neurospheres/well was determined following 10 days for ten different wells. Spheres that contained more than 20 cells were scored and the results are presented as percentages of
maximal neurospheres formed compared to control cells28,29. IN VITRO LIMITING DILUTION ASSAY For the in vitro limiting dilution assay, GSCs were plated in 96-well plates in decreasing
numbers of cells (50, 20, 10, 5, 2, and 1) per well. Following 10 days, the number of spheres was determined for each well. Extreme limiting dilution was analyzed as recently reported32.
REAL-TIME POLYMERASE CHAIN REACTION (RT-PCR) Total RNA was extracted using RNeasy midi kit according to the manufacturer’s instructions (Qiagen, Frederick, MD). Reverse transcription
reaction was carried out using 2-μg total RNA as previously described28,31. Briefly, reactions were run on an ABI Prism 7000 Sequence Detection System (Applied Biosystems, Foster City, CA).
Cycle threshold (Ct) values were obtained from the ABI 7000 software. S12 levels were used as controls. The primer sequences are described in Table S2. WESTERN BLOT ANALYSIS Cell pellet
preparation and Western Blot analyses were performed as previously described28,29,30. TRANSWELL MIGRATION ASSAY Transwell chambers (BD Biosciences, San Jose, CA) were used for analyzing cell
migration as recently reported29,34 In brief, cells (25,000/well) were incubated for 3 h in culture medium with 10% fetal bovine serum in the bottom chambers. The total number of the
migrated cells was determined in fixed and stained cells (0.05% crystal violet for 5 min). CELL VIABILITY ASSAY Cells were washed with phosphate-buffered saline (PBS), centrifugated in 3000
rpm for 5 min and the cell pellet was incubated in PBS containing 0.4% trypan to stain the dead cells. The number of Trypan-blue stained cells was determined using a Countess II FL (Thermo
Fisher, MA, USA). ISOLATION OF GSC-DERIVED EVS EVs were prepared as previously described31,35 using sequential centrifugation at 300 × _g_ for 10 min, 2500 × _g_ for 20 min, 10,000 × _g_ for
30 min and 110,000 × _g_ for 90 min. The pellet was then resuspended in PBS and washed twice followed by filtration using a 0.22-μm filter. The protein content of the enriched EV fractions
was determined using the Micro BCA assay kit (ThermoFischer Scientific, Oregon City, OR). The expression of the exosome markers CD63, CD81, and CD9 was analyzed by Western blot and the
quantification of the isolated EVs was performed using the ExoELISA-Ultra CD63 kit according to the manufacturer’s instructions. For the exosome treatment, 0.5 × 108 EVs were added to the
cultured cells. IMAGESTREAMX ANALYSIS Microglial cells were treated with GSC-derived EVs labeled with CellTracker Red (ThermoFisher, Waltham, MA) for 24 h. Cells were excited using 561-nm
laser, and cell fluorescence of approximately 104 cells per sample was captured and photographed using an ImageStreamX high-resolution imaging flow cytometer (Amnis Co., Seattle, WA) as
previously described35. The samples were gated to obtain a population of captured single-cell images of living cells, then gated for the cells in focus using the gradient root mean square
feature. Cells incubated with or without labeled EVs were compared for the intensity of the red channel fluorescence. Images were analyzed using IDEAS 6.0 software (Amnis Co., Seattle, WA).
MIR-504 REPORTER For analyzing miR-504 delivery, a miR-504 luciferase reporter plasmid was employed as previously described for miR-12436. A unique miR-504 binding site, which is a fully
complementary sequence of mature miR-504, was cloned downstream of luciferase reporter gene of the pMiR-Luc reporter vector from Signosis, Inc. (Santa Clara, CA). For the mCherry reporter,
the luciferase gene of pMiR-Luc reporter vector was replaced with mCherry-N1 obtained from Clontech (Mountain View, CA). PHAGOCYTOSIS ANALYSIS Human microglial cells were plated alone or in
coculture with GSCs. Phagocytosis was determined using the pHrodo™ Green zymosan bioparticle assay (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. Briefly,
microglia plated alone and in the presence of GSCs were incubated with a solution of pHrodo Green zymosan bioparticles in Live Cell Imaging Solution (0.5 mg/ml). Phagocytosis was determined
after 120 min using fluorescence plate reader at Ex/Em 509/533. MIRNA ARRAY PROCESSING AND ANALYSIS All experiments were performed using Affymetrix HU GENE1.0st oligonucleotide arrays and
GeneChip miRNA 4.0 Array (ThermoFisher). Sample processing was performed according to the protocol provided by the company. The rest of the analysis was performed using Partek® Genomics
SuiteTM software, version 6.6 (©2012 Partek, Inc.). miRNA data were summarized using RMA and standardized by sketch-quantile normalization. Differential expression was performed via ANOVA.
Significant miRNAs were selected to have at least 1.5-fold change and a _P_ value < 0.05. Results were visualized by volcano plot. Functional analysis was conducted by Ingenuity software
using the core analysis on differential miRNA lists. The panel of measured miRNAs (a list of all measured miRNAs) was used as the background set for enrichment tests. Networks included up to
35 miRNAs and mRNAs. TCGA DATA ANALYSIS Expression data were downloaded for TCGA cases from the Broad Firehose portal (http://gdac.broadinstitute.org/). GBM cases were assayed by microarray
for miRNA expression6. The level 3, batch-adjusted, expression data file captured mature miRNA quantification (file date: 12/10/2014). Low-grade glioma (LGG) cases were assayed by
miRNA-sequencing37. The level 3 data file contained expression data per mature miRNA as reads per million miRNAs mapped (file date: 12/10/2014). GBM and LGG cases were assayed by
mRNA-sequencing5. The level 3 data file contained RSEM normalized data38, quantified per-gene as the normalized count (file date: 12/10/2014). Expression data are used continuously,
discretized by quantile, or dichotomized at the median as high/low as appropriate for the research question. Clinical data and molecular classifications were taken from the recent
publication of the TCGA Glioma Analysis Working Group6. Comparison of mean expression between groups was performed by one-way ANOVA followed by Tukey’s corrected two-sample tests, which
adjust for multiple comparisons to maintain the family-wise error rate. XENOGRAFT STUDIES Following the guidelines of Henry Ford Hospital’s Institutional Animal Care and Use Committee,
dissociated GSCs (3 × 105 cells) transduced with a lentivirus vector expressing a control pre-miR or pre-miR-504 were inoculated intracranially into nude mice (Nu/Nu) as previously
described32. Briefly, animals were anesthetized and injected with the GSCs through a 3-mm hole to the right of the bregma at a depth of 2.5 mm and a rate of 5 μL/30 s. All animals were
monitored daily and sacrificed at the first signs of neurological deficit. STATISTICAL ANALYSIS The results are presented as the mean values ± SD. The data of patient specimens are presented
graphically with median and interquartile range noted. Data were analyzed using ANOVA or a Student’s _t_ test with correction for data sets with unequal variances. Data were analyzed on a
log 2 scale as appropriate. Kaplan–Meier estimates of the survival time from diagnosis until death or last follow-up were used for outcome analysis. Differences in survival curves between
groups were assessed by the log-rank test. Cox regression was used to construct multivariable models of survival including miRNA expression, age at diagnosis, IDH mutation status and grade.
RESULTS FUNCTIONAL CLUSTERING AND NETWORKS ASSOCIATED WITH MIRNAS THAT DISTINGUISH GSCS FROM HNSCS To define the patterns of miRNA expressions that are unique to GSCs and associated with
their tumorigenicity and mesenchymal characteristics, we used a miRNA array chip for 12 GSCs and three different cultures of hNSCs. We first compared the miRNA expression of GSCs and hNSCs.
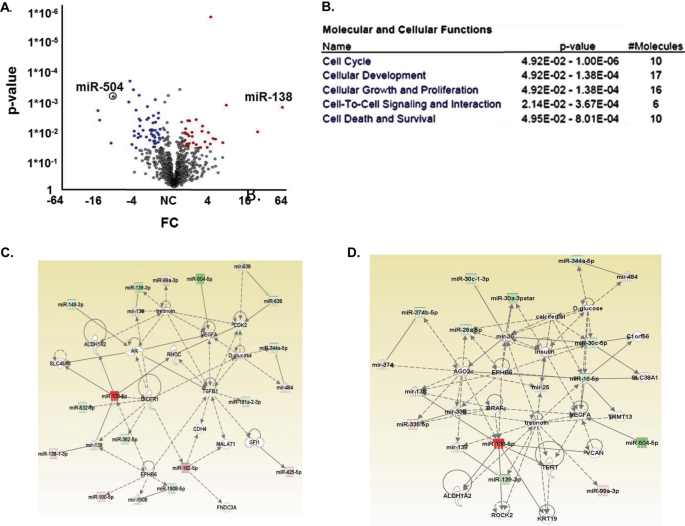
miRNAs were identified using cutoffs for ≥1.5-fold differential expression and a significant _P_ value (_P_ ≤ 0.05), as listed in Table S3 and as shown in a volcano plot (Fig. 1A). Thirty
miRNAs were significantly upregulated, and 55 miRNAs were downregulated in GSCs relative to hNSCs. These miRNAs were further analyzed by functional enrichment and network analysis using
Ingenuity Pathway Analysis (IPA; Ingenuity Systems, http://www.ingenuity.com). IPA analysis identified clusters of miRNAs that are associated with well-known oncogenic pathways including
cell cycle, cellular development, cellular growth and proliferation, cell-to-cell signaling and interaction and cell death and survival (Fig. 1B). IPA was also used to generate three
networks of altered miRNA interactions consisting of at least 15 miRNAs from the miRNA lists (Figs. 1C, S1A, B). These networks are associated with miRNA biogenesis including regulation of
Dicer1 and AGO2 (Figs. 1C, S1A) and of oncogenes such as TERT, MYC, CASP2, CASP10, BCL2, and TP73 (Fig. S1a). An important oncogenic pathway that was also identified is associated with
increased regulation of classical EMT mediators such as Smad2/3, Smad6/7, TGFβ1, and Dicer (Figs. 1C, S1B). We then identified miRNAs that were expressed in GSCs but not in hNSCs and found
that 10 miRNAs were significantly upregulated in GSCs while 37 were downregulated (Fig. S1D). The IPA of these miRNA clusters generated two networks, each containing more than 10 miRNAs
(Figs. 1D, S1C). These networks consist of proteins related to cell survival (p53 and TERT) and miRNA biogenesis (Dicer and AGO2) similar to the pathways that were obtained in the initial
comparison of the GSCs and hNSCs (Fig. 1D). In addition, the two key mesenchymal markers ZEB2 and RUNX1 were also identified in these networks (Fig. S1C). MIR-504 IS DOWNREGULATED IN GBMS
AND GSCS Using RT-PCR analysis we first validated some of the miR array results (Figs. 2A, S2). Since miR-504 was one of the most downregulated miRNAs in GSCs compared with hNSCs (Fig. 1A),
we focused on this miRNA as a potential inhibitor of the tumorigenicity of GSCs. The expression of miR-504 in GBM specimens was also significantly increased in normal brain compared with
astrocytic tumor specimens (Fig. 2B). We then analyzed the relative expression of miR-504 in the different subtypes of GBM using TCGA6. There are 339 GBM cases in our study that have
IDH/Methylation subtyping. This subtyping splits the IDHmut-noncodel class into two groups according to methylation pattern (Fig. 2C). The G-CMIP-low class has a lower level of methylation
globally and has been found to have worse prognosis. The IDHwt GBM tumors are split into three groups. Two align with expression class, as mesenchymal-like and classic-like, and the third
has a distinct methylation pattern, denoted as LGm6–GBM (Fig. 2C, ANOVA, _P_ < 0.0001). Gray lines indicate significant difference between groups (post hoc _t_ test, _P_ < 0.05).
Analysis of miR-504 expression in LLGs demonstrated that the expression of miR-504 was higher on average in grade II glioma compared with grade III (Fig. 2D). There are 509 LGG cases in our
study that have IDH/methylation subtyping. This subtyping splits the IDHmut class into three groups according to methylation pattern (Fig. 2E). The G-CMIP-high class has a highest miR-504
expression. The IDHwt LGG tumors are also split into three groups. The classic-like and PA-like are two groups with the lowest and highest expression, respectively. While the
mesenchymal-like group has intermediate expression (Fig. 2E). Gray lines indicate significant difference between groups (post hoc _t_ test, _P_ < 0.05). While survival differences were
observed by IDH-mutation status (data not shown), there was no evidence that miR-504 expression has independent prognostic value beyond the two new WHO 2016 diagnostic groups, GBM with IDHwt
and GBM with IDH mutant. A Kaplan–Meier plot demonstrates that the quarter of patients with lowest expression of miR-504 has the worst survival outcome among grade II and III glioma (Fig.
2F; log-rank _P_ = 0.00136 overall; log-rank _P_ = 0.00402, 0.00123, 0.0111 Q1 vs. Q2–4, respectively). Collectively, these results indicate that miR-504 expression is inversely correlated
with tumor aggressiveness and poor prognosis. MIR-504 INHIBITS THE STEMNESS AND MESENCHYMAL TRANSIT OF GSCS To examine the effect of miR-504 we overexpressed it in GSCs (Fig. 3SA) and found
that pre-miR-504 markedly decreased the expression of the stemness markers Oct4 and Nanog and increased the expression of the astrocytic marker GFAP in both GSC-1 and GSC-2 (Fig. 3A). The
role of miR-504 in the stemness of GSCs was further examined on the self-renewal ability and neurosphere formation of these cells. Overexpression of miR-504 in GSCs decreased their ability
to form neurospheres as indicated by analyzing secondary neurosphere formation (Fig. 3B), the extreme limiting dilution assay (Fig. 3C) and spheroid size (Fig. 3D), suggesting that miR-504
inhibited the stemness potential of GSCs and increased their differentiation. miR-504 overexpression did not induce cell death in the GSCs as determined by trypan blue staining (data not
shown). In addition, overexpression of miR-504 also decreased GSC migration (Fig. 3E) and the expression of the mesenchymal markers CTGF, fibronectin 1 (FN), and YKL-40 (Fig. 3F). These
results demonstrate that miR-504 acts as a negative regulator of the stemness, mesenchymal phenotypes, and migration of GSCs. We further examined the effects of miR-504 overexpression on the
tumorigenic capacity of GSCs in vivo. For these experiments we employed GSC-1 transduced with lentivirus vectors expressing pre-miR-504. Intracranial implantation of these cells into
immunocompromised mice resulted in significantly smaller xenografts (4.35 ± 1.33 mm2; _n_ = 5) compared with those expressing a control pre-miR (11.96 ± 4.20 mm2; _N_ = 5, _P_ < 0.005)
(Fig. 3G). In addition, overexpression of miR-504 increased the mean survival of tumor-bearing mice compared with the control miR group (66 days vs. 42 days, _P_ < 0.0001, _N_ = 11/group)
(Fig. 3H). These data demonstrate that overexpression of miR-504 in GSCs decreased their tumorigenicity which is in line with the favorable prognosis of patients with tumors that express
high levels of this miR. GRB10 IS A TARGET OF MIR-504 AND MEDIATES THE INHIBITORY EFFECTS OF THIS MIR ON GSCS Bioinformatics analysis identified Grb10 as a potential target of miR-504 which
was also recently reported as a validated one39. Using the Grb10 3′-UTR- tagged to luciferase, we demonstrated a direct targeting of Grb10 by miR-504 (Fig. 4A). We then analyzed the
expression of Grb10 in hNSCs and GSCs (_N_ = 10) and found that this gene was highly expressed in GSCs compared to hNSCs (Fig. 4B). Using TCGA we analyzed the expression of Grb10 in various
subtypes of glioma and found that the expression of Grb10 was significantly higher in GBM compared with low-grade tumors as determined by histology criteria (Fig. 4C) and the WHO grade (Fig.
4D). In addition, analysis of methylation glioma subtypes demonstrated that Grb10 was highly expressed in the mesenchymal subtype and exhibited lowest expression in the G-CIMP high tumors
(Fig. 4E). Kaplan–Meier analysis (Fig. 4F) shows that the quarter of patients with highest expression of Grb10 have the worst survival outcomes (log-rank _P_ = 6.39e−12 overall; log-rank _P_
= 5.13e−11, 6.15e−07, 2e−05, Q4 vs. Q1–3, respectively). Dichotomizing miR-504 at the 25th and Grb10 at the 75th percentiles, demonstrates that the low/high pattern of miR-504 and Grb10,
respectively, more clearly identifies a subset of patients with poor overall survival compared to each expression alone (Fig. 4G, log-rank _P_ = 2.67e−8 overall; log-rank _P_ = 0.0012,
2.89e−12, 0.0003, low/high vs. high/high, high/low and low/low, respectively). We then examined the role of Grb10 in GSC functions and demonstrated that its overexpression in GSCs increased
(Fig. 4H), while its silencing decreased (Fig. 4H) the stemness and mesenchymal phenotypes of GSCs, similar to the effects of miR-504. Overexpression of a Grb10 plasmid lacking 3′-UTR
induced a modest upregulation of self-renewal of the GSCs and partially abrogated the inhibitory effect of miR-504 on the mesenchymal phenotype (Fig. 4I) and the self-renewal (Fig. 4J) of
the treated GSCs. These results demonstrate that targeting Grb10 by miR-504 mediates at least in part the inhibitory effects of miR-504 on the stemness and mesenchymal phenotypes of GSCs.
OVEREXPRESSION OF MIR-504 IN GSCS PROMOTES M1 (PRO-INFLAMMATORY) PHENOTYPES OF COCULTURED MICROGLIAL CELLS Glioma cells and GSCs have been demonstrated to induce polarization of microglia
toward the M2 phenotype (anti-inflammatory/pro-tumorigenic) by the secretion of cytokines and EV-derived miRNAs23,40,41. In addition, we recently reported that EVs can deliver exogenous
miRNAs both in vitro and in vivo31,36. We therefore hypothesized that the overexpressed miR-504 in GSCs can be transferred to neighboring cells and therefore affects not only the oncogenic
functions of the GSCs but also their interactions with neighboring cells such as microglia. For these studies we employed co-cultures of microglial cells and GSCs overexpressing pre-miR-504
or a control pre-miR in transwell plates with 1-μm filters that allow only the transfer of soluble factors and EVs as described in Fig. 5A. Coculturing of microglia with control GSCs or
cells transduced with lentivirus expressing a control pre-miR (Fig. 5A, D) resulted in a relative increased expression of the M2-associated markers, CD209 and TGF-β and in the decreased
expression of CD86 and TNF-α (Fig. 5B, E). GSC coculturing also increased the phagocytosis of microglia cells (Fig. 5C). In contrast, transduction of the GSCs with pre-miR-504 (Fig. 5D)
decreased the expression of the M2-associated markers, increased the expression of CD86 and TNF-α (Fig. 5E) and decreased phagocytosis (Fig. 5F). To verify that miR-504 was delivered by the
GSCs to the cocultured microglial cells, we employed a miR-504 reporter tagged to luciferase that allows the quantification of the transferred miRNA as was previously reported33. Microglial
cells were transduced with a lentivirus vector expressing the miR-504-luciferase reporter and the GSCs were transduced with lentivirus vector expressing either control pre-miR or pre-miR-504
(Fig. 5D). As presented in Fig. 5G, microglia that were co-cultured with GSCs overexpressing pre-miR-504 exhibited decreased luciferase activity indicating that miR-504 was transferred by
the cocultured GSCs. These results indicate that miR-504 was transferred by GSCs to cocultured microglial cells. We further analyzed the expression of miR-504 in the cocultured microglial
cells and found that they expressed high levels of this miR compared with microglia cocultured with GSCs expressing a control miR (Fig. 5H). Finally, we demonstrated that overexpression of
miR-504 in microglial cells upregulated the relative expression of the M1 markers CD86 and TNF-α (Fig. 5I). EV-ASSOCIATED MIR-504 DERIVED FROM GSCS INDUCES MICROGLIA M1 PHENOTYPES
EV-associated miRNAs are implicated in the cross-talk of GSCs and microglia33,35. To determine the role of EVs in the delivery of miR-504 to microglial cells, we first analyzed the
expression of miR-504 in EVs derived from GSC-1 that were transduced with lentivirus vectors expressing control pre-miR or pre-miR-504. EVs were isolated from GSC-1 cultures using
differential ultracentrifugation as previously described35 and were analyzed for the expression of CD63, CD81, and CD9 (Fig. S3B). The amount of the secreted EVs was determined using ELISA
of CD63 antibodies and was found to be comparable in GSC-1 overexpressing con-miR or miR-504 (data not shown). We found that EVs isolated from GSC-1 overexpressing miR-504 expressed
significantly higher levels of miR-504 compared with EVs isolated from GSC-1 expressing a control pre-miR (Figs. 6A, S3C). We next demonstrated the transfer of EVs from GSC-1 to microglial
cells using ImageStreamX analysis. In these studies, GSC-1 derived EVs labeled with CellTracker Red were added to microglial cells and their fluorescence was determined 12 h later. The EVs
were efficiently internalized and accumulated in the microglial cells (Fig. 6B). Incubation of microglial cells that express the miR-504 reporter with EVs that were isolated from GSC-1
expressing either the con-miR or miR-504 (Fig. 6C) demonstrated the functional delivery of the miR-504 to the microglial cells (Fig. 6D), which resulted in decreased expression of CD209 and
TGF-β, and increased expression of CD86 and TNF-α (Fig. 6E), similar to our observations with cocultured GSCs (Fig. 5D). To demonstrate that the delivered miR-504 mediated the effects of the
GSC-1 derived EVs, we examined the effects of EVs isolated from GSC-1 overexpressing miR-504 on the differentiation of microglial cells transfected with a miR-504 antagomir (Fig. 6F) and
found that these effects were markedly abrogated, whereas, no significant inhibition was observed in microglia transfected with a control antagomir (Fig. 6G). These results indicate that the
transfer of miR-504 by GSC-derived EVs mediated, at least partly, the increased M1 phenotypes of the microglial cells induced by the cocultured GSCs. DISCUSSION GBM is one of the most
aggressive, infiltrative and incurable tumors with an average patient survival of around 14–16 months1,2. GBM therapy resistance and recurrence are primarily attributed to the existence of
GSCs8,9,10. Therefore, targeting these cells is an essential component of any successful therapeutic approach. Recent studies demonstrated that the mesenchymal differentiation of GBM is
associated with acquisition of stemness characteristics, tumor aggressiveness, therapy resistance, and poor clinical outcome16,17,18,42. Thus, deciphering the mechanisms underlying the
mesenchymal differentiation of GSCs is essential for identifying novel therapeutic targets and improving patient survival. miRNAs have emerged as attractive therapeutic targets due to their
critical roles in the regulation of major cell processes such as cell proliferation, stemness, and apoptosis that are key components in cancer initiation and progression29,30,31,43. In
addition, specific miRNAs have been implicated in controlling the mesenchymal differentiation of tumor cells26,44,45. Using miRNA microarray analysis of GSCs in comparison to hNSCs, we
identified 85 miRNAs that were significantly altered in the GSCs compared with hNSCs. These miRNAs were found to be associated with well-known tumorigenic pathways including cell cycle,
cellular development, cellular growth and proliferation, cell-to-cell signaling and interaction, and cell death and survival. These findings indicate that alterations in miRNA expression are
associated with deregulation in pathways which contribute to the tumorigenic phenotypes of GSCs. Additional differences in miRNA expression between GSCs and hNSCs were also associated with
the EMT process and included pathways regulating Smad2/3, Smad6/7 TGFβ146, and Dicer47, suggesting that the expression of specific miRNAs in GSCs regulate their own expression in parallel
with the tumorigenic characteristics of these cells. Finally, other IPA networks identified ZEB2 and RUNX1, two major regulators of the EMT pathway7, as mainly enriched in GSCs. One of the
most downregulated miRNAs in GSCs compared to hNSCs was miR-504. We further found that miR-504 expression was significantly lower in GBM as compared to normal brains and exhibited a
grade-dependent expression. In addition, using the TCGA portal, we found that miR-504 expression was significantly increased in the G-CIMP high glioma and more generally in IDH-mutant GBM,
whereas, it was considerably lower in the IDH-wt glioma classes including the mesenchymal-like subtypes. In agreement with the lower expression of miR-504 in more high grade tumors and in
the mesenchymal subtype, we found that overexpression of miR-504 inhibited the self-renewal and mesenchymal phenotypes of GSCs, Collectively, the current results highlight miR-504 as a
potential tumor suppressor miRNA and as a negative regulator of the tumorigenicity of GBM and GSCs. Our results of a role of miR-504 as a tumor suppressor in glioma and as an inhibitor of
mesenchymal transformation are in agreement with recent publications48,49,50,51,52. The current studies present new data regarding the expression of miR-504 in patient-derived GSCs compared
with NSCs and in specific subtypes of gliomas including patient survival data. In addition, the current studies focus on the functions of miR-504 in GSCs including their tumorgenicity using
intracranial xenografts. The role of miR-504 in oncogenic processes appears to be tumor dependent. Thus, in gastric cancer miR-504 expression was decreased by the tumor suppressor gene
Trefoil factor 1 (TFF1) that leads to the activation of p5353, whereas miR-504 had a dual function in oral squamous cell carcinoma54. Moreover, serum expression of miR-504 were demonstrated
to differentiate between primary and metastatic brain tumors55, suggesting a role of this miR as a diagnostic marker and a mediator of the interaction of glioma and non-CNS cells. The
inhibitory effects of miR-504 on GSCs were at least partly mediated by Grb10, which was recently reported as a validated miR-504 target in vascular endothelial cells39. Grb10 is an imprinted
gene that is differentially expressed from two promoters and in the brain it is paternally expressed56. The role of Grb10 in tumorigenesis is just beginning to be understood57,58. Using
TCGA analysis, we demonstrated that Grb10 was highly expressed in more aggressive glioma tumors and its expression was directly correlated with worse prognosis. In addition, overexpression
of Grb10 in GSCs promoted their aggressiveness, whereas silencing exerted an opposite effect and abrogated the inhibitory effect of miR-504. Thus, our studies demonstrated the miR-504/Grb10
pathway as an important regulator of the stemness-EMT process in GSCs. Additional targets of miR-504 were reported in glioma and a recent study reported that miR-504 inhibited EMT by
targeting the Frizzled-7-mediated the Wnt-β-catenin pathway51. GSCs have been reported to promote the differentiation of microglia toward the M2/anti-inflammatory phenotype, an effect that
is more pronounced in mesenchymal GSCs23,40,41. The M2 microglia phenotype in turn acts to support the migration and aggressiveness of the tumor cells and inhibition of anti-tumor immune
response24,59,60. The cross talk between microglia and GSCs is mediated by secreted cytokines and EV-derived miRNAs23,40,41,61 In view of our recent reports that EVs can also deliver
exogenous miRNA to neighboring cells31,36, we hypothesized that overexpression of miR-504 in GSCs may be transferred to neighboring cells via EVs. Using a miR-504 reporter that can directly
detect changes in miRNA levels, we found that GSCs overexpressing miR-504 delivered this miRNA to cocultured microglia via EVs. We also found that the increased relative expression of M2
markers induced by GSCs was abrogated in microglia co-cultured with GSCs overexpressing a miR-504 mimic. The mechanisms by which the EV-derived miR-504 exerts its effects on the microglial
cells are currently being studied. Since miR-504 decreased the stemness and mesenchymal differentiation of GSCs, it is possible that other factors or miRNAs that are secreted by the
transduced GSCs can also contribute to the induction of this change in the microglial phenotype. The mutual crosstalk between glioma cells and microglia highlights the importance of soluble
factors as potential therapeutic targets. Indeed, EV-associated miRNAs have been reported to play an important role in intercellular interactions in both physiological and pathological
conditions62,63. In addition, overexpressed miRNA mimics have been also reported to be delivered by EVs to neighboring cells36,39. Our data indicate that overexpressing miR-504 in GSCs
affects not only the functions of the tumor cells but also the tumor-promoting activity of microglia and probably macrophages in the tumor microenvironment, thereby amplifying the
therapeutic effect of miR-504 (Fig. 7). CONCLUSIONS The acquisition of mesenchymal phenotypes has been associated with increased stemness, infiltration, and aggressive phenotypes in GSCs64.
Therefore, identifying therapeutic targets that can interfere with this process is of utmost importance. Performing comparative analyses of hNSCs and GSCs, we identified novel miRNAs and
potential target networks that are associated with the stemness and mesenchymal transit of GSCs. miR-504 is downregulated in GSCs and exerts inhibitory effects on the functions of these
cells via the targeting of Grb10 that acts as an oncogene in GBM and GSCs. Importantly, the overexpression of miR-504 in GSCs not only inhibits the tumorigenic potential of GSCs in vitro and
in vivo but can be also transferred to microglial calls and promote their M1 polarization. Thus, the antitumor effects of RNA-based therapy in cancer cells can further exert a bystander
effect on the tumor microenvironment via EV delivery. DATA AVAILABILITY Data are available in the Supplementary files. REFERENCES * Furnari, F. B. et al. Malignant astrocytic glioma:
genetics, biology, and paths to treatment. _Genes Dev._ 21, 2683–2710 (2007). Article CAS PubMed Google Scholar * Desjardins, A. et al. Chemotherapy and novel therapeutic approaches in
malignant gliomas. _Front. Biosci._ 10, 2645–2668 (2005). Article CAS PubMed Google Scholar * Phillips, H. S. et al. Molecular subclasses of high-grade glioma predict prognosis,
delineate a pattern of disease progression, and resemble stages in neurogenesis. _Cancer Cell_ 9, 157–173 (2006). Article CAS PubMed Google Scholar * Verhaak, R. G. W. et al. Integrated
genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. _Cancer Cell_ 17, 98–110 (2010). Article CAS PubMed
PubMed Central Google Scholar * Ceccarelli, M. et al. Molecular profiling reveals biologically discrete subsets and pathways of progression in diffuse glioma. _Cell_ 164, 550–563 (2016).
Article CAS PubMed PubMed Central Google Scholar * Brennan, C. W. et al. The somatic genomic landscape of glioblastoma. _Cell_ 155, 462–477 (2013). Article CAS PubMed PubMed Central
Google Scholar * Carro, M. S. et al. The transcriptional network for mesenchymal transformation of brain tumours. _Nature_ 463, 318–325 (2010). Article CAS PubMed Google Scholar *
Singh, S. K. et al. Identification of a cancer stem cell in human brain tumors. _Cancer Res._ 63, 5821–5828 (2003). CAS PubMed Google Scholar * Vescovi, A. L., Galli, R. & Reynolds,
B. A. Brain tumour stem cells. _Nat. Rev. Cancer_ 6, 425–436 (2006). Article CAS PubMed Google Scholar * Lathia, J. D., Mack, S. C., Mulkearns-Hubert, E. E., Valentim, C. L. L. &
Rich, J. N. Cancer stem cells in glioblastoma. _Genes Dev._ 29, 1203–1217 (2015). Article CAS PubMed PubMed Central Google Scholar * Lim, D. A. et al. Relationship of glioblastoma
multiforme to neural stem cell regions predicts invasive and multifocal tumor phenotype. _Neuro. Oncol._ 9, 424–429 (2007). Article PubMed PubMed Central Google Scholar * Matarredona, E.
R. & Pastor, A. M. Neural stem cells of the subventricular zone as the origin of human glioblastoma stem cells. Therapeutic implications. _Front. Oncol._ 9, 779–824 (2019). Article
PubMed PubMed Central Google Scholar * Nieto, M. A., Huang, R. Y. Y. J., Jackson, R. A. A. & Thiery, J. P. P. EMT: 2016. _Cell_ 166, 21–45 (2016). Article CAS PubMed Google Scholar
* Tso, C. L. et al. Primary glioblastomas express mesenchymal stem-like properties. _Mol. Cancer Res._ 4, 607–619 (2006). Article CAS PubMed Google Scholar * Behnan, J., Finocchiaro,
G. & Hanna, G. The landscape of the mesenchymal signature in brain tumours. _Brain_ 142, 847–866 (2019). Article PubMed PubMed Central Google Scholar * Fedele, M., Cerchia, L.,
Pegoraro, S., Sgarra, R. & Manfioletti, G. Proneural-mesenchymal transition: Phenotypic plasticity to acquire multitherapy resistance in glioblastoma. _Int. J. Mol. Sci._ 20, 2746–2760
(2019). Article CAS PubMed Central Google Scholar * Halliday, J. et al. In vivo radiation response of proneural glioma characterized by protective p53 transcriptional program and
proneural-mesenchymal shift. _Proc. Natl Acad. Sci. USA_ 111, 5248–5253 (2014). Article CAS PubMed PubMed Central Google Scholar * Bhat, K. P. L. et al. Mesenchymal differentiation
mediated by NF-κB promotes radiation resistance in glioblastoma. _Cancer Cell_ 24, 331–346 (2013). Article CAS PubMed Google Scholar * Quail, D. F. & Joyce, J. A. The
microenvironmental landscape of brain tumors. _Cancer Cell_ 31, 326–341 (2017). Article CAS PubMed PubMed Central Google Scholar * Zanotto-Filho, A. et al. Inflammatory landscape of
human brain tumors reveals an NFκB dependent cytokine pathway associated with mesenchymal glioblastoma. _Cancer Lett._ 390, 176–187 (2017). Article CAS PubMed Google Scholar * Wang, Q.
et al. Tumor evolution of glioma intrinsic gene expression subtype associates with immunological changes in the microenvironment. _Cancer Cell_ 32, 42–56 (2017). Article PubMed CAS PubMed
Central Google Scholar * Chen, Z. & Hambardzumyan, D. Immune microenvironment in glioblastoma subtypes. _Front. Immunol._ 9, 1004–1012 (2018). Article PubMed CAS PubMed Central
Google Scholar * Abels, E. R. et al. Glioblastoma-associated microglia reprogramming is mediated by functional transfer of extracellular miR-21. _Cell Rep._ 28, 3105–3119.e7 (2019). Article
CAS PubMed PubMed Central Google Scholar * Hambardzumyan, D., Gutmann, D. H. & Kettenmann, H. The role of microglia and macrophages in glioma maintenance and progression. _Nat.
Neurosci._ 19, 20–27 (2015). Article CAS Google Scholar * Godlewski, J., Krichevsky, A. M., Johnson, M. D., Chiocca, E. A. & Bronisz, A. Belonging to a network-microRNAs,
extracellular vesicles, and the glioblastoma microenvironment. _Neuro Oncol._ 17, 652–662 (2015). Article CAS PubMed Google Scholar * Møller, H. G. et al. A systematic review of MicroRNA
in glioblastoma multiforme: Micro-modulators in the mesenchymal mode of migration and invasion. _Mol. Neurobiol._ 47, 131–144 (2013). Article PubMed CAS Google Scholar * Liu, C. &
Tang, D. G. MicroRNA regulation of cancer stem cells. _Cancer Res._ 71, 5950–5954 (2011). Article CAS PubMed PubMed Central Google Scholar * Bier, A. et al. MicroRNA-137 is
downregulated in glioblastoma and inhibits the stemness of glioma stem cells by targeting RTVP-1. _Oncotarget_ 4, 665–676 (2013). Article PubMed PubMed Central Google Scholar * Lee, H.
K. et al. MicroRNA-145 is downregulated in glial tumors and regulates glioma cell migration by targeting connective tissue growth factor. _PLoS ONE_ 8, e54652 (2013). Article CAS PubMed
PubMed Central Google Scholar * Giladi, N. D. et al. RTVP-1 promotes mesenchymal transformation of glioma via a STAT-3/IL-6-dependent positive feedback loop. _Oncotarget_ 6, 22680–22697
(2015). Article PubMed PubMed Central Google Scholar * Lee, H. K. et al. Mesenchymal stem cells deliver synthetic microRNA mimics to glioma cells and glioma stem cells and inhibit their
cell migration and self-renewal. _Oncotarget_ 4, 346–361 (2013). Article PubMed PubMed Central Google Scholar * Jiang, W. et al. Repurposing phenformin for the targeting of glioma stem
cells and the treatment of glioblastoma. _Oncotarget_ 35, 56456–56470 (2016). Article Google Scholar * Quartararo, C. E., Reznik, E., deCarvalho, A. C., Mikkelsen, T. & Stockwell, B.
R. High-throughput screening of patient-derived cultures reveals potential for precision medicine in glioblastoma. _ACS Med. Chem. Lett._ 6, 948–952 (2015). Article CAS PubMed PubMed
Central Google Scholar * Ziv-Av, A. et al. RTVP-1 regulates glioma cell migration and invasion via interaction with N-WASP and hnRNPK. _Oncotarget_ 6, 19826–19840 (2015). Article PubMed
PubMed Central Google Scholar * Bier, A. et al. Placenta-derived mesenchymal stromal cells and their exosomes exert therapeutic effects in Duchenne muscular dystrophy. _Biomaterials_ 174,
67–78 (2018). Article CAS PubMed Google Scholar * Lee, H. K., Finniss, S., Cazacu, S., Xiang, C. & Brodie, C. Mesenchymal stem cells deliver exogenous miRNAs to neural cells and
induce their differentiation and glutamate transporter expression. _Stem Cells Dev._ 23, 2851–2861 (2014). Article CAS PubMed Google Scholar * Brat, D. J. et al. Comprehensive,
integrative genomic analysis of diffuse lower-grade gliomas. _N. Engl. J. Med._ 372, 2481–2498 (2015). Article CAS PubMed Google Scholar * Li, B. & Dewey, C. N. RSEM: accurate
transcript quantification from RNA-Seq data with or without a reference genome. _BMC Bioinform._ 12, 323–339 (2011). Article CAS Google Scholar * Reddy, M. A. et al. Regulation of
vascular smooth muscle cell dysfunction under diabetic conditions by MIR-504. _Arterioscler. Thromb. Vasc. Biol._ 36, 864–873 (2016). Article CAS PubMed PubMed Central Google Scholar *
De Vrij, J. et al. Glioblastoma-derived extracellular vesicles modify the phenotype of monocytic cells. _Int. J. Cancer_ 137, 1630–1642 (2015). Article PubMed CAS Google Scholar * Van
Der Vos, K. E. et al. Directly visualized glioblastoma-derived extracellular vesicles transfer RNA to microglia/macrophages in the brain. _Neuro Oncol._ 18, 58–69 (2016). Article PubMed
CAS Google Scholar * Li, G. et al. ALDH1A3 induces mesenchymal differentiation and serves as a predictor for survival in glioblastoma. _Cell Death Dis._ 9, 1190–1201 (2018). Article CAS
PubMed PubMed Central Google Scholar * Rynkeviciene, R. et al. Non-coding RNAs in glioma. _Cancers_ 22, 11–46 (2018). Google Scholar * Tang, J. et al. Molecular mechanisms of microRNAs
in regulating epithelial-mesenchymal transitions in human cancers. _Cancer Lett._ 371, 301–313 (2016). Article CAS PubMed Google Scholar * Hao, J. et al. MicroRNA control of
epithelial-mesenchymal transition in cancer stem cells. _Int. J. Cancer_ 135, 1019–1027 (2014). Article CAS PubMed Google Scholar * Han, J., Alvarez-Breckenridge, C. A., Wang, Q. E.
& Yu, J. TGF-β signaling and its targeting for glioma treatment. _Am. J. Cancer Res._ 5, 945–955 (2015). CAS PubMed PubMed Central Google Scholar * Martello, G. et al. A microRNA
targeting dicer for metastasis control. _Cell_ 141, 1195–1207 (2010). Article CAS PubMed Google Scholar * Guan, Y. et al. Downregulation of microRNA-504 is associated with poor prognosis
in high-grade glioma. _Int. J. Clin. Exp. Pathol._ 8, 727–734 (2015). CAS PubMed PubMed Central Google Scholar * Cui, R., Guan, Y., Sun, C., Chen, L., Bao, Y., Li, G., Qiu, B., Meng,
X., Pang, C. & Wang, Y. A tumor-suppressive microRNA, miR-504, inhibits cell proliferation and promotes apoptosis by targeting FOXP1 in human glioma. _Cancer Lett._ 28, 1–11 (2016).
Article CAS Google Scholar * Seo, J., Jin, D., Choi, C. H. & Lee, H. Integration of microRNA, mRNA, and protein expression data for the identification of cancer-related microRNAs.
_Plos ONE_ 5, e0168412 (2017). Article CAS Google Scholar * Liu, Q., Guan, Y., Li, Z., Wang, Y., Liu, Y., Cui, R. & Wang, Y. miR-504 suppresses mesenchymal phenotype of glioblastoma
by directly targeting the FZD7-mediated Wnt-β-catenin pathway. _Exp. Clin. Cancer Res._ 16, 358–376 (2019). Article CAS Google Scholar * Ma, X., Yoshimoto, K., Guan, Y., Hata, N.,
Mizoguchi, M., Sagata, N., Murata, H., Kuga, D., Amano, T., Nakamizo, A. & Sasaki, T. Associations between microRNA expression and mesenchymal marker gene expression in glioblastoma.
_Neuro Oncol._ 14, 1153–1162 (2012). Article CAS PubMed PubMed Central Google Scholar * Soutto, M. et al. TFF1 activates p53 through down-regulation of miR-504 in gastric cancer.
_Oncotarget_ 5, 5663–5673 (2014). Article PubMed PubMed Central Google Scholar * Yang, M. H. et al. Connective tissue growth factor modulates oral squamous cell carcinoma invasion by
activating a miR-504/FOXP1 signalling. _Oncogene_ 31, 2401–2411 (2012). Article CAS PubMed Google Scholar * Jin, Z., Jin, R. H., Ma, C., Li, H. S. & Xu, H. Y. Serum expression level
of miR-504 can differentiate between glioblastoma multiforme and solitary brain metastasis of non-small cell lung carcinoma. _J. BUON_ 22, 474–480 (2017). PubMed Google Scholar *
Plasschaert, R. N. & Bartolomei, M. S. Tissue-specific regulation and function of Grb10 during growth and neuronal commitment. _Proc. Natl Acad. Sci. USA_ 112, 6841–6947 (2015). Article
CAS PubMed Google Scholar * Khan, M. I. et al. Proproliferative function of adaptor protein GRB10 in prostate carcinoma. _FASEB J._ 33, 3198–3211 (2019). Article CAS PubMed Google
Scholar * Mroue, R., Huang, B., Braunstein, S., Firestone, A. J. & Nakamura, J. L. Monoallelic loss of the imprinted gene Grb10 promotes tumor formation in irradiated Nf1+/- mice. _PLoS
Genet._ 11, 1–25 (2015). Google Scholar * Wu, A. et al. Glioma cancer stem cells induce immunosuppressive macrophages/microglia. _Neuro Oncol._ 12, 1113–1125 (2010). Article CAS PubMed
PubMed Central Google Scholar * Li, W. & Graeber, M. B. The molecular profile of microglia under the influence of glioma. _Neuro Oncol._ 14, 958–978 (2012). Article CAS PubMed
PubMed Central Google Scholar * Bronisz, A., Godlewski, J. & Chiocca, E. A. Extracellular vesicles and microRNAs: their role in tumorigenicity and therapy for brain tumors. _Cell. Mol.
Neurobiol._ 36, 361–376 (2016). Article CAS PubMed PubMed Central Google Scholar * Fan, Q. et al. The emerging role of exosome-derived non-coding RNAs in cancer biology. _Cancer Lett._
414, 107–115 (2018). Article CAS PubMed Google Scholar * Nakano, I., Garnier, D., Minata, M. & Rak, J. Extracellular vesicles in the biology of brain tumour stem cells -
Implications for inter-cellular communication, therapy and biomarker development. _Semin. Cell Dev. Biol._ 40, 17–26 (2015). Article CAS PubMed Google Scholar * Iser, I. C., Pereira, M.
B., Lenz, G. & Wink, M. R. The epithelial-to-mesenchymal transition like process in glioblastoma: an updated systematic review and in silico investigation. _Med. Res. Rev._ 37, 271–313
(2017). Article CAS PubMed Google Scholar Download references ACKNOWLEDGEMENTS We thank Dr. Hae Kyung Lee and Susan Finniss for their technical support and Susan McPhee for her diligent
editing of the manuscript. This work was supported by the William and Karen Davidson Fund, Hermelin Brain Tumor Center and by ICRF (C.B.). AUTHOR INFORMATION Author notes * These authors
contributed equally: Ariel Bier, Xin Hong, Simona Cazacu AUTHORS AND AFFILIATIONS * The Mina and Everard Goodman Faculty of Life Sciences, Bar-Ilan University, Ramat-Gan, Israel Ariel Bier,
Hodaya Goldstein, Daniel Rand, Hiba Waldman Ben-Asher, Aharon Brodie & Chaya Brodie * Davidson Laboratory of Cell Signaling and Tumorigenesis, Hermelin Brain Tumor Center, Department of
Neurosurgery, Henry Ford Hospital, Detroit, MI, USA Xin Hong, Simona Cazacu, Cunli Xiang, Wei Jiang & Chaya Brodie * Department of Neurosurgery, Sheba Medical Center, Henry Ford
Hospital, Detroit, MI, USA Moshe Attia * Department of Public Health Sciences, Henry Ford Hospital, Detroit, MI, USA Ruicong She & Laila M. Poisson Authors * Ariel Bier View author
publications You can also search for this author inPubMed Google Scholar * Xin Hong View author publications You can also search for this author inPubMed Google Scholar * Simona Cazacu View
author publications You can also search for this author inPubMed Google Scholar * Hodaya Goldstein View author publications You can also search for this author inPubMed Google Scholar *
Daniel Rand View author publications You can also search for this author inPubMed Google Scholar * Cunli Xiang View author publications You can also search for this author inPubMed Google
Scholar * Wei Jiang View author publications You can also search for this author inPubMed Google Scholar * Hiba Waldman Ben-Asher View author publications You can also search for this author
inPubMed Google Scholar * Moshe Attia View author publications You can also search for this author inPubMed Google Scholar * Aharon Brodie View author publications You can also search for
this author inPubMed Google Scholar * Ruicong She View author publications You can also search for this author inPubMed Google Scholar * Laila M. Poisson View author publications You can
also search for this author inPubMed Google Scholar * Chaya Brodie View author publications You can also search for this author inPubMed Google Scholar CORRESPONDING AUTHOR Correspondence to
Chaya Brodie. ETHICS DECLARATIONS ETHICS APPROVAL AND CONSENT TO PARTICIPATE All human materials were used in accordance with the policies of the institutional review board at Henry Ford
Hospital, Detroit, MI. CONFLICT OF INTEREST The authors declare that they have no conflict of interest. ADDITIONAL INFORMATION PUBLISHER’S NOTE Springer Nature remains neutral with regard to
jurisdictional claims in published maps and institutional affiliations. Edited by G. Calin SUPPLEMENTARY INFORMATION SUPPLEMENTARY FIGURE S1 SUPPLEMENTARY FIGURE S2 SUPPLEMENTARY FIGURE S3
SUPPLEMENTARY FIGURE LEGENDS SUPPLEMENTARY TABLES RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative Commons Attribution 4.0 International License, which permits
use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the
Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated
otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds
the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/. Reprints and
permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Bier, A., Hong, X., Cazacu, S. _et al._ miR-504 modulates the stemness and mesenchymal transition of glioma stem cells and their interaction
with microglia via delivery by extracellular vesicles. _Cell Death Dis_ 11, 899 (2020). https://doi.org/10.1038/s41419-020-03088-3 Download citation * Received: 05 December 2019 * Revised:
13 August 2020 * Accepted: 24 August 2020 * Published: 22 October 2020 * DOI: https://doi.org/10.1038/s41419-020-03088-3 SHARE THIS ARTICLE Anyone you share the following link with will be
able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing
initiative