- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT mRNA localization is an essential mechanism of gene regulation and is required for processes such as stem-cell division, embryogenesis and neuronal plasticity. It is not known which
features in the _cis_-acting mRNA localization elements (LEs) are specifically recognized by motor-containing transport complexes. To the best of our knowledge, no high-resolution structure
is available for any LE in complex with its cognate protein complex. Using X-ray crystallography and complementary techniques, we carried out a detailed assessment of an LE of the _ASH1_
mRNA from yeast, its complex with its shuttling RNA-binding protein She2p, and its highly specific, cytoplasmic complex with She3p. Although the RNA alone formed a flexible stem loop, She2p
binding induced marked conformational changes. However, only joining by the unstructured She3p resulted in specific RNA recognition. The notable RNA rearrangements and joint action of a
globular and an unfolded RNA-binding protein offer unprecedented insights into the step-wise maturation of an mRNA-transport complex. Access through your institution Buy or subscribe This is
a preview of subscription content, access via your institution ACCESS OPTIONS Access through your institution Access Nature and 54 other Nature Portfolio journals Get Nature+, our
best-value online-access subscription $29.99 / 30 days cancel any time Learn more Subscribe to this journal Receive 12 print issues and online access $209.00 per year only $17.42 per issue
Learn more Buy this article * Purchase on SpringerLink * Instant access to full article PDF Buy now Prices may be subject to local taxes which are calculated during checkout ADDITIONAL
ACCESS OPTIONS: * Log in * Learn about institutional subscriptions * Read our FAQs * Contact customer support SIMILAR CONTENT BEING VIEWED BY OTHERS MULTIPLE PARA/MIND ATPASES COORDINATE THE
POSITIONING OF DISPARATE CARGOS IN A BACTERIAL CELL Article Open access 05 June 2023 STRUCTURE OF FULL-LENGTH ERGIC-53 IN COMPLEX WITH MCFD2 FOR CARGO TRANSPORT Article Open access 16 March
2024 STRUCTURAL ASYMMETRY GOVERNS THE ASSEMBLY AND GTPASE ACTIVITY OF MCRBC RESTRICTION COMPLEXES Article Open access 20 November 2020 ACCESSION CODES PRIMARY ACCESSIONS PROTEIN DATA BANK *
5M0H * 5M0I * 5M0J REFERENCED ACCESSIONS PROTEIN DATA BANK * 1XLY * 4FNJ * 4WNL REFERENCES * Holt, C.E. & Bullock, S.L. Subcellular mRNA localization in animal cells and why it matters.
_Science_ 326, 1212–1216 (2009). Article CAS PubMed PubMed Central Google Scholar * Tolino, M., Köhrmann, M. & Kiebler, M.A. RNA-binding proteins involved in RNA localization and
their implications in neuronal diseases. _Eur. J. Neurosci._ 35, 1818–1836 (2012). Article PubMed Google Scholar * St Johnston, D. Moving messages: the intracellular localization of
mRNAs. _Nat. Rev. Mol. Cell Biol._ 6, 363–375 (2005). Article CAS PubMed Google Scholar * Marchand, V., Gaspar, I. & Ephrussi, A. An intracellular transmission control protocol:
assembly and transport of ribonucleoprotein complexes. _Curr. Opin. Cell Biol._ 24, 202–210 (2012). Article CAS PubMed Google Scholar * Buxbaum, A.R., Haimovich, G. & Singer, R.H. In
the right place at the right time: visualizing and understanding mRNA localization. _Nat. Rev. Mol. Cell Biol._ 16, 95–109 (2015). Article CAS PubMed Google Scholar * Munro, T.P. et al.
Mutational analysis of a heterogeneous nuclear ribonucleoprotein A2 response element for RNA trafficking. _J. Biol. Chem._ 274, 34389–34395 (1999). Article CAS PubMed Google Scholar *
Chao, J.A. et al. ZBP1 recognition of beta-actin zipcode induces RNA looping. _Genes Dev._ 24, 148–158 (2010). Article CAS PubMed PubMed Central Google Scholar * Patel, V.L. et al.
Spatial arrangement of an RNA zipcode identifies mRNAs under post-transcriptional control. _Genes Dev._ 26, 43–53 (2012). Article CAS PubMed PubMed Central Google Scholar * Jambhekar,
A. & Derisi, J.L. Cis-acting determinants of asymmetric, cytoplasmic RNA transport. _RNA_ 13, 625–642 (2007). Article CAS PubMed PubMed Central Google Scholar * Pratt, C.A. &
Mowry, K.L. Taking a cellular road-trip: mRNA transport and anchoring. _Curr. Opin. Cell Biol._ 25, 99–106 (2013). Article CAS PubMed Google Scholar * Bullock, S.L., Ringel, I.,
Ish-Horowicz, D. & Lukavsky, P.J. A′-form RNA helices are required for cytoplasmic mRNA transport in _Drosophila_. _Nat. Struct. Mol. Biol._ 17, 703–709 (2010). Article CAS PubMed
PubMed Central Google Scholar * Simon, B., Masiewicz, P., Ephrussi, A. & Carlomagno, T. The structure of the SOLE element of oskar mRNA. _RNA_ 21, 1444–1453 (2015). Article CAS
PubMed PubMed Central Google Scholar * Bertrand, E. et al. Localization of _ASH1_ mRNA particles in living yeast. _Mol. Cell_ 2, 437–445 (1998). Article CAS PubMed Google Scholar *
Gonzalez, I., Buonomo, S.B., Nasmyth, K. & von Ahsen, U. _ASH1_ mRNA localization in yeast involves multiple secondary structural elements and Ash1 protein translation. _Curr. Biol._ 9,
337–340 (1999). Article CAS PubMed Google Scholar * Müller, M. et al. A cytoplasmic complex mediates specific mRNA recognition and localization in yeast. _PLoS Biol._ 9, e1000611 (2011).
Article PubMed PubMed Central CAS Google Scholar * Olivier, C. et al. Identification of a conserved RNA motif essential for She2p recognition and mRNA localization to the yeast bud.
_Mol. Cell. Biol._ 25, 4752–4766 (2005). Article CAS PubMed PubMed Central Google Scholar * Jambhekar, A. et al. Unbiased selection of localization elements reveals cis-acting
determinants of mRNA bud localization in _Saccharomyces cerevisiae_. _Proc. Natl. Acad. Sci. USA_ 102, 18005–18010 (2005). Article CAS PubMed PubMed Central Google Scholar * Niedner,
A., Edelmann, F.T. & Niessing, D. Of social molecules: the interactive assembly of ASH1 mRNA-transport complexes in yeast. _RNA Biol._ 11, 998–1009 (2014). Article PubMed PubMed
Central Google Scholar * Edelmann, F.T., Niedner, A. & Niessing, D. ASH1 mRNP-core factors form stable complexes in absence of cargo RNA at physiological conditions. _RNA Biol._ 12,
233–237 (2015). Article PubMed PubMed Central Google Scholar * Heym, R.G. et al. _In vitro_ reconstitution of an mRNA-transport complex reveals mechanisms of assembly and motor
activation. _J. Cell Biol._ 203, 971–984 (2013). Article CAS PubMed PubMed Central Google Scholar * Sladewski, T.E., Bookwalter, C.S., Hong, M.S. & Trybus, K.M. Single-molecule
reconstitution of mRNA transport by a class V myosin. _Nat. Struct. Mol. Biol._ 20, 952–957 (2013). Article CAS PubMed PubMed Central Google Scholar * Böhl, F., Kruse, C., Frank, A.,
Ferring, D. & Jansen, R.P. She2p, a novel RNA-binding protein tethers _ASH1_ mRNA to the Myo4p myosin motor via She3p. _EMBO J._ 19, 5514–5524 (2000). Article PubMed PubMed Central
Google Scholar * Long, R.M., Gu, W., Lorimer, E., Singer, R.H. & Chartrand, P. She2p is a novel RNA-binding protein that recruits the Myo4p-She3p complex to _ASH1_ mRNA. _EMBO J._ 19,
6592–6601 (2000). Article CAS PubMed PubMed Central Google Scholar * Müller, M. et al. Formation of She2p tetramers is required for mRNA binding, mRNP assembly, and localization. _RNA_
15, 2002–2012 (2009). Article PubMed PubMed Central CAS Google Scholar * Shahbabian, K., Jeronimo, C., Forget, A., Robert, F. & Chartrand, P. Co-transcriptional recruitment of Puf6
by She2 couples translational repression to mRNA localization. _Nucleic Acids Res._ 42, 8692–8704 (2014). Article CAS PubMed PubMed Central Google Scholar * Shen, Z., Paquin, N.,
Forget, A. & Chartrand, P. Nuclear shuttling of She2p couples ASH1 mRNA localization to its translational repression by recruiting Loc1p and Puf6p. _Mol. Biol. Cell_ 20, 2265–2275
(2009). Article CAS PubMed PubMed Central Google Scholar * Niedner, A., Müller, M., Moorthy, B.T., Jansen, R.P. & Niessing, D. Role of Loc1p in assembly and reorganization of
nuclear ASH1 messenger ribonucleoprotein particles in yeast. _Proc. Natl. Acad. Sci. USA_ 110, E5049–E5058 (2013). Article CAS PubMed PubMed Central Google Scholar * Takizawa, P.A.
& Vale, R.D. The myosin motor, Myo4p, binds _Ash1_ mRNA via the adapter protein, She3p. _Proc. Natl. Acad. Sci. USA_ 97, 5273–5278 (2000). Article CAS PubMed PubMed Central Google
Scholar * Bobola, N., Jansen, R.P., Shin, T.H. & Nasmyth, K. Asymmetric accumulation of Ash1p in postanaphase nuclei depends on a myosin and restricts yeast mating-type switching to
mother cells. _Cell_ 84, 699–709 (1996). Article CAS PubMed Google Scholar * Chartrand, P., Meng, X.H., Hüttelmaier, S., Donato, D. & Singer, R.H. Asymmetric sorting of ash1p in
yeast results from inhibition of translation by localization elements in the mRNA. _Mol. Cell_ 10, 1319–1330 (2002). Article CAS PubMed Google Scholar * Long, R.M. et al. Mating type
switching in yeast controlled by asymmetric localization of _ASH1_ mRNA. _Science_ 277, 383–387 (1997). Article CAS PubMed Google Scholar * Takizawa, P.A., Sil, A., Swedlow, J.R.,
Herskowitz, I. & Vale, R.D. Actin-dependent localization of an RNA encoding a cell-fate determinant in yeast. _Nature_ 389, 90–93 (1997). Article CAS PubMed Google Scholar *
Niessing, D., Hüttelmaier, S., Zenklusen, D., Singer, R.H. & Burley, S.K. She2p is a novel RNA binding protein with a basic helical hairpin motif. _Cell_ 119, 491–502 (2004). Article
CAS PubMed Google Scholar * Singh, N., Blobel, G. & Shi, H. Hooking She3p onto She2p for myosin-mediated cytoplasmic mRNA transport. _Proc. Natl. Acad. Sci. USA_ 112, 142–147 (2015).
Article CAS PubMed Google Scholar * Tijerina, P., Mohr, S. & Russell, R. DMS footprinting of structured RNAs and RNA-protein complexes. _Nat. Protoc._ 2, 2608–2623 (2007). Article
CAS PubMed PubMed Central Google Scholar * Ferré-D'Amaré, A.R., Zhou, K. & Doudna, J.A. A general module for RNA crystallization. _J. Mol. Biol._ 279, 621–631 (1998). Article
PubMed Google Scholar * Gonsalvez, G.B. et al. RNA-protein interactions promote asymmetric sorting of the _ASH1_ mRNA ribonucleoprotein complex. _RNA_ 9, 1383–1399 (2003). Article CAS
PubMed PubMed Central Google Scholar * Lunde, B.M., Moore, C. & Varani, G. RNA-binding proteins: modular design for efficient function. _Nat. Rev. Mol. Cell Biol._ 8, 479–490 (2007).
Article CAS PubMed PubMed Central Google Scholar * Shen, Z., St-Denis, A. & Chartrand, P. Co-transcriptional recruitment of She2p by RNA pol II elongation factor Spt4-Spt5/DSIF
promotes mRNA localization to the yeast bud. _Genes Dev._ 24, 1914–1926 (2010). Article CAS PubMed PubMed Central Google Scholar * Calabretta, S. & Richard, S. Emerging roles of
disordered sequences in RNA-binding proteins. _Trends Biochem. Sci._ 40, 662–672 (2015). Article CAS PubMed Google Scholar * McGinnis, J.L. et al. In-cell SHAPE reveals that free 30S
ribosome subunits are in the inactive state. _Proc. Natl. Acad. Sci. USA_ 112, 2425–2430 (2015). Article CAS PubMed PubMed Central Google Scholar * Fürtig, B., Nozinovic, S., Reining,
A. & Schwalbe, H. Multiple conformational states of riboswitches fine-tune gene regulation. _Curr. Opin. Struct. Biol._ 30, 112–124 (2015). Article PubMed CAS Google Scholar * Chen,
W. & Moore, M.J. The spliceosome: disorder and dynamics defined. _Curr. Opin. Struct. Biol._ 24, 141–149 (2014). Article CAS PubMed Google Scholar * Lange, S. et al. Simultaneous
transport of different localized mRNA species revealed by live-cell imaging. _Traffic_ 9, 1256–1267 (2008). Article CAS PubMed Google Scholar * Edelmann, F.T., Niedner, A. &
Niessing, D. Production of pure and functional RNA for in vitro reconstitution experiments. _Methods_ 65, 333–341 (2014). Article CAS PubMed Google Scholar * Janowski, R. et al. Roquin
recognizes a non-canonical hexaloop structure in the 3′-UTR of Ox40. _Nat. Commun._ 7, 11032 (2016). Article CAS PubMed PubMed Central Google Scholar * Zuker, M. Mfold web server for
nucleic acid folding and hybridization prediction. _Nucleic Acids Res._ 31, 3406–3415 (2003). Article CAS PubMed PubMed Central Google Scholar * Petoukhov, M.V. et al. New developments
in the ATSAS program package for small-angle scattering data analysis. _J. Appl. Crystallogr._ 45, 342–350 (2012). Article CAS PubMed PubMed Central Google Scholar * Winn, M.D. et al.
Overview of the CCP4 suite and current developments. _Acta Crystallogr. D Biol. Crystallogr._ 67, 235–242 (2011). Article CAS PubMed PubMed Central Google Scholar * Morin, A. et al.
Collaboration gets the most out of software. _eLife_ 2, e01456 (2013). Article PubMed PubMed Central Google Scholar * Joosten, R.P., Long, F., Murshudov, G.N. & Perrakis, A. The
PDB_REDO server for macromolecular structure model optimization. _IUCrJ_ 1, 213–220 (2014). Article CAS PubMed PubMed Central Google Scholar * Du, T.G. et al. Nuclear transit of the
RNA-binding protein She2 is required for translational control of localized ASH1 mRNA. _EMBO Rep._ 9, 781–787 (2008). Article CAS PubMed PubMed Central Google Scholar Download
references ACKNOWLEDGEMENTS We thank V. Roman for her support, and M. Seiler and M. Feldbrügge for helpful discussions. We acknowledge the use of the X-ray crystallography platform of the
Helmholtz Zentrum München, NMR measurements at the Bavarian NMR center and SAXS measurements at the facility of the SFB1035, Technische Universität München. This work was supported by the
Deutsche Forschungsgemeinschaft (SFB1035 to M.S.; SPP1935 to M.S. and D.N.; FOR2333 to R.-P.J. and D.N.; SFB646 to D.N.) and by the Bayerisch-Französisches Hochschulzentrum (BFHZ) to D.N.
AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * Institute of Structural Biology, Helmholtz Zentrum München – German Research Center for Environmental Health, Neuherberg, Germany Franziska
Theresia Edelmann, Roland Gerhard Heym, Robert Janowski, Michael Sattler & Dierk Niessing * Department Chemie, Center for Integrated Protein Science Munich at Biomolecular NMR
Spectroscopy, Technische Universität München, Garching, Germany Andreas Schlundt, Ralf Stehle & Michael Sattler * Interfaculty Institute of Biochemistry, University of Tübingen,
Tübingen, Germany Andreas Jenner, Muhammad Ibrahim Syed & Ralf-Peter Jansen * Department of Cell Biology, Biomedical Center of the Ludwig-Maximilians-Universität München,
Planegg-Martinsried, Germany Annika Niedner-Boblenz & Dierk Niessing * Architecture et Réactivité de l'ARN, Université de Strasbourg, Centre National de la Recherche Scientifique,
Institut de Biologie Moléculaire et Cellulaire, Strasbourg, France Jean-Christophe Paillart Authors * Franziska Theresia Edelmann View author publications You can also search for this author
inPubMed Google Scholar * Andreas Schlundt View author publications You can also search for this author inPubMed Google Scholar * Roland Gerhard Heym View author publications You can also
search for this author inPubMed Google Scholar * Andreas Jenner View author publications You can also search for this author inPubMed Google Scholar * Annika Niedner-Boblenz View author
publications You can also search for this author inPubMed Google Scholar * Muhammad Ibrahim Syed View author publications You can also search for this author inPubMed Google Scholar *
Jean-Christophe Paillart View author publications You can also search for this author inPubMed Google Scholar * Ralf Stehle View author publications You can also search for this author
inPubMed Google Scholar * Robert Janowski View author publications You can also search for this author inPubMed Google Scholar * Michael Sattler View author publications You can also search
for this author inPubMed Google Scholar * Ralf-Peter Jansen View author publications You can also search for this author inPubMed Google Scholar * Dierk Niessing View author publications You
can also search for this author inPubMed Google Scholar CONTRIBUTIONS F.T.E., A.S., R.G.H., A.J., A.N.-B., M.I.S. and R.S. conducted the experiments. F.T.E., A.S., R.G.H., R.-P.J., J.-C.P.
and D.N. designed the experiments. F.T.E., A.S., R.G.H., A.N.-B., R.S., R.J., M.S., R.-P.J. and D.N. analyzed the data. F.T.E. and D.N. wrote the paper. CORRESPONDING AUTHOR Correspondence
to Dierk Niessing. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing financial interests. INTEGRATED SUPPLEMENTARY INFORMATION SUPPLEMENTARY FIGURE 1 ASSESSMENT OF
SYNERGISTIC RNA RECOGNITION BY THE SHE2P-SHE3P COMPLEX IN ELECTROPHORETIC MOBILITY SHIFT ASSAYS (EMSA). (A) E3-(51 nt) LE was previously shown to mediate full synergistic She2p-She3p binding
and is incorporated into motile particles (Heym, R.G. _et al_., _J Cell Biol_. 203, 971-84 (2013); Müller, M. _et al_., _PLoS Biol_. 9, e1000611 (2011)). The single-stranded regions at the
5’ and 3’ ends of E3 (51 nt) relate to a large bulge in the context of full-length _ASH1_ mRNA (Fig. 1a). Deletion of either the 5’end or part of te 3’ regions in the E3-(51 nt) mutants Δ1
and Δ2, respectively, resulted in wild-type binding. In contrast, a complete deletion of the single-stranded bases at the 3’ end abolished binding (Δ3). The results indicate that bases
1812-1814 are indispensable. Final She2p concentrations were 0.02 μM, 0.06 μM, 0.18 μM, 0.54 μM, 1.61 μM and 4.86 μM. (B) A deletion of the upper part of the stem and of the nona-loop (nt
1786-1802) combined with an insertion of a more compact tetra-loop (38 nt-loop) still allowed for high-affinity binding by She2p and She3p. She2p concentrations were 0.01 μM, 0.03 μM, 0.10
μM, 0.30 μM, 0.90 μM and 2.70 μM. (c) Comparative EMSAs of She2p wild type and a cystein-mutated, N-terminally truncated She2p version (She2p(6-246, C-S)). The latter shows wild type-like
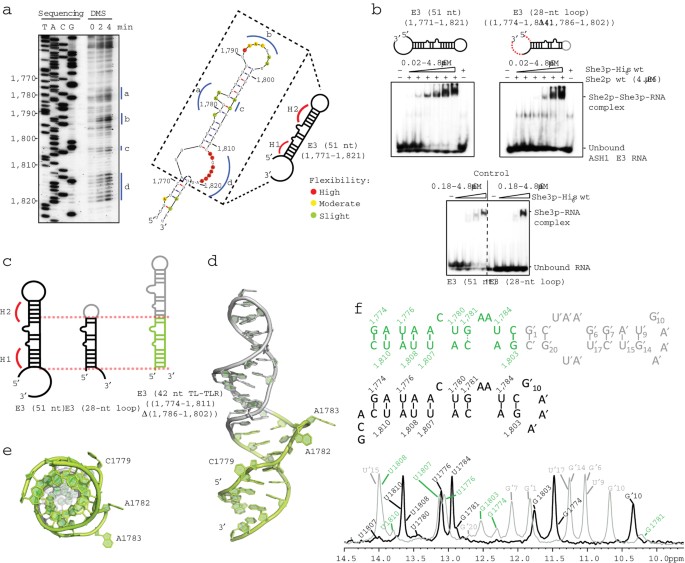
ternary complex formation with She3p and the E3-(51 nt) RNA. Each EMSA is representative for three independent experiments. SUPPLEMENTARY FIGURE 2 NMR ANALYSIS OF THE FREE RNA IN SOLUTION.
(A) Agarose gel with the denatured RNA sample before crystallization and after extraction from crystals indicates no degradation, suggesting that the lack of electron density in this region
is due to structural flexibility. (B) 2D-imino NOESY spectra of _ASH1_-E3 (28 nt-loop) and E3-(42 nt TL-TLR) RNAs showing the sequentially assigned observable imino protons. Resonance labels
are color-coded as indicated in Fig. 1f. Solid lines in the scheme represent unambiguous hydrogen-bonds as obtained from the sequential assignments. The broken line for G1781-C1805 in the
42-mer indicates an assignment of the G1781 imino proton inferred by exclusion, although no cross peak to adjacent nucleotides was observed. Overall the spectra are consistent with the
existence of the base pairing observed in the crystal structure (Fig. 1d,e) also in solution. However, significant line broadening of imino resonances and undetectable imino signals in the
bulged region between H1 and H2 indicate dynamics and flexibility in this part of the RNA. Bases marked with apostrophe indicate heterologous GAAA tetra-loop containing sequences, not
belonging to the E3 LE. Bases highlighted with red boxes show peak broadening in the 1D imino spectrum indicating dynamics. SUPPLEMENTARY FIGURE 3 ASSESSMENT OF SHE2P-RNA STRUCTURE. (A-E)
She2p binding of She3p(382-405) is not physiologic. Native crystals of She2p with RNA diffracted best when soaked with a short She3p peptide (residues 382-405). (A) In the She2p-RNA
co-structure (Fig. 2a) additional electron density was observed that could not be assigned to She2p or RNA. The location of this not well-ordered electron density, however, overlaps with a
previously reported interaction site of She3p (364-368) (Singh, N. _et al_., _Proc Natl Acad Sci U S A_. 112, 142-7 (2015)). (B) The corresponding residues from fragment 382-405 were modeled
into electron density (398-405, green) and compared to the previously published peptide 363-368 (red). Interactions were similar but not identical. (C) A sequence comparison of the
previously described interaction motifs of She3p(364-368) (grey box) and of She3p(382-405) revealed similarities. (D) Crystal structure of She2p(6-246, C-S) with the _ASH1_-E3 (28 nt-loop)
RNA at 2.41 Å resolution in front view and rotated 90° around the vertical axis. Non-physiologically bound She3p fragments are depicted in green. (E) Isothermal titration calorimetry (ITC)
experiments with She2p and She3p(382-405) (top) showed no binding even at final She3p concentrations of 157 μM in three independent experiments. Since the positive control (She3p residues
364-368) (Singh, N. _et al_., _Proc Natl Acad Sci U S A_. 112, 142-7 (2015)) showed binding to She2p (bottom) in two independent experiments, we conclude that the electron density of
She3p(382-405) in the co-structure with She2p and RNA is non-physiologic. (F) Close-up of interactions in the kinked region of E3 RNA in the She2p-bound state. Watson-Crick base pairings,
hydrogen bonding, base stacking, and water interactions are shown. For better visualization, She2p is hidden. (G) Superposition of the two RNA chains E3 (pink) and E3’ (grey) observed in the
She2p-bound form. The sole difference is visible at the very 3’ base (RMSD of all atoms = 0.5 Å). SUPPLEMENTARY FIGURE 4 COMPARISON OF FREE RNA WITH SHE2P-BOUND RNA IN SOLUTION BY SMALL
ANGLE X-RAY SCATTERING. (a) Overlay of the RNA models from the E3-(42 nt-TL-TLR) RNA crystal structure (green), where the heterologous GAAA-donor acceptor has been replaced _in silico_ by a
shorter GAAA tetra-loop (grey), and of the E3 (28 nt-loop) RNA from the She2p-She3p bound complex (magenta). Strong rearrangements in the RNA secondary structure pinpoint to a large
conformational change upon complex formation (see also Fig. 2c). (b-d) Different RNA models (red) are fitted against the scattering curve of E3-(28 nt-loop) RNA recorded at 1.5 mg/ml
concentration in single experiments. (b) The calculated scattering curves for both the bound (in (c): pink, kinked) and unbound (in c: green, elongated) RNAs, assuming that both
conformations could be present (with fitted fractions kinked RNA: elongated RNA = 0.72: 0.27) does not fit well to the scattering curve especially at low (0.03 Å-1) and high (0.3 Å-1) q
values, which is also reflected by the high χ2 value of 7.6. (c) When testing a kinked and a melted, single-stranded RNA model (ssRNA) (depicted in blue; ratio kinked RNA: ssRNA = 0.74:
0.26) the fit becomes better and χ2 decrases to 3.9. (d) Assuming that an elongated and a single-stranded species (ratio elongated RNA: ssRNA = 0.65: 0.34) exists in solution, the χ2 value
improves to 1.7. (e) The most accurate fit could be achieved by fitting a combination of the kinked, single-stranded and elongated RNA (ratio kinked: ssRNA: elongated = 0.12: 0.33: 0.55).
Here χ2 was 1.6. SAXS data support the observation that the E3-(28 nt-loop) RNA has an elongated conformation in solution. The fact that a single-stranded species is needed to fit the data
suggests an ensemble of RNA conformations. Notably, the elongated form represents a major population consistent with the SAXS and NMR data. SUPPLEMENTARY FIGURE 5 CONTROL EMSA AND SURFACE
REPRESENTATION OF THE TERNARY COMPLEX. (A) Control EMSA related to Fig. 3b showing that none of the used She3p truncation mutants (331-405 or 331-425) is able to bind E3-(28 nt-loop) RNA in
the absence of She2p at the given experimental concentrations. (B) Schematic drawing of She2p(6-246, C-S) and the N-terminally fused flexible linkers consisting of five (GGSGG) or ten amino
acids (GGSGG)2. (C) EMSA with She2p-She3p fusion constructs. Since She2p and She3p interact with a 1:1 ratio, they also form synergistic complexes when fused to each other with a
glycine-serine linker with different length. The last lane on the right side contains She2p(6-246, C-S) fused to the (GGSGG)2 linker as a control, not showing any band shift. (D) She2p alone
is depicted in surface representation with its contact sites for She3p (green) and RNA (magenta) in front view and 90 ° rotated around the vertical axis. (E) The electrostatic surface
potential of She2p shows that E3-(28 nt-loop) RNA (magenta) binds at the positively charged area (blue) in the middle of the She2p tetramer, whereas negatively charged residues (red)
surround the binding region. She3p is depicted in green. (F) Stereo view of the She2p-She3p complex with _ASH1_ E3 (28 nt-loop) RNA. (G) Ternary complex as depicted in Fig. 3b with She2p in
surface representation. Red area marks She2p amino acids 164-179 that were previously shown to UV crosslink with E3 (51 nt) (Müller, M. _et al_., _PLoS Biol_. 9, e1000611 (2011)). Dashed
blue line shows the anticipated projection of the RNA over the crosslinking site. This depicted single-stranded loop region followed by a double-stranded stem (right side) is consistent with
the experimentally validated secondary structure shown in Fig. 1a. EMSAs in A and C are representatives for three independent experiments each. SUPPLEMENTARY FIGURE 6 REPRESENTATIVE EMSAS
FOR APPARENT KD DETERMINATION. (A-D) Binding affinities of E3 variants that showed ternary complex formation in Fig. 4 were quantified in EMSA experiments. Respective control EMSAs show
unspecific RNA binding of She3p at high protein concentrations. Nevertheless, when She2p is additionally present, band intensities of the shifted complex increase, thus reflecting the
specific ternary complex of She2p-She3p and RNA. In (A) radioactively labeled E3-(51 nt) RNA was used. In (B) E3 (28 nt-loop) was assessed. (C) and (D) show EMSAs where RNA mutants “M1” and
“M2” were tested. (E) Table summarizing apparent mean KD values ± s.d. for _ASH1_ E3-RNA mutants in complex with She2p and She3p. KDs were calculated from three independent experiments using
the one-site binding equation. While E3 (51 nt) forms the ternary complex with a KD of 0.20 ± 0.03 μM, the minimal E3-(28 nt-loop) RNA bound She2p and She3p just slightly weaker. Replacing
U1780 by the pyrimidine C in mutant “M2” decreased the affinity to a KD of 0.44 ± 0.09 μM, whereas mutating U1780 to purine A in mutant “M1” had a more severe effect with a KD of 1.05 ± 0.33
μM. (F) Representative EMSA of the crystallized She2p(6-246, C-S)-(GGSGG)2-She3p(331-405) fusion protein with _ASH1_ E3-(28 nt-loop) RNA shows high affinity binding. The apparent KD was 112
± 29 nM. Square brackets marked by an asterisk delimit the area of the gel, which was used for quantification in A-D and F. All EMSA were performed in three independent experiments.
SUPPLEMENTARY FIGURE 7 ANALYSES OF SHE2P MUTANTS AND FUSION CONSTRUCTS WITH SHE3P. (A) EMSA testing _ASH1_-E3 (51-nt) RNA-binding by different She2p-She3p fusion constructs. She2p(6-246,
C-S) fused to the (GGSGG)2 linker and to She3p(331-405) shows strong ternary complex formation. When in She3p K340, R341 and Y345 are mutated to alanines, band shifts occur only at higher
protein concentrations, indicating a reduced RNA-binding affinity. The fusion protein only containing She3p amino acids 331 to 346 is still able to form a weak ternary complex, confirming
that this region constitutes an important interaction site. The last lane on the right side contains She2p(6-246, C-S) fused to the ten amino acid glycine-serine linker (GGSGG)2 without
She3p as a control, not showing any band shift. (B) Control EMSA related to Fig. 4d. Unless stated otherwise 4.86 μM protein(s) were tested for _ASH1_-E3 (51-nt) RNA-binding. (C) CD spectra
of She2p wild type in comparison to the double mutant She2p(E172A, F176A) confirmed the α-helical composition of both protein variants and the secondary structure integrity of the mutant.
EMSAs in A and B are representatives for three independent experiments each. CD spectra in C were recorded once. SUPPLEMENTARY FIGURE 8 REPRESENTATIVE EMSAS OF SHE3P MUTANTS FOR KD
CALCULATIONS. (A) His6-SUMO-She3p(331-405) and selected single-amino acid mutants were tested with She2p and _ASH1_-E3 (51 nt) for their ternary complex-formation. Since distinct
She2p-She3p-RNA complexes could be detected for wild-type She3p(331-405) and She3p(331-405; R342A) apparent mean KD values ± s.d. were determined. KDs were calculated from three independent
experiments using the one-site binding equation. Square brackets marked by an asterisk delimit the area of the gel, which was used for quantification. His6-SUMO-tagged proteins were used
because they yielded higher amounts of protein after purification. (B) Table summarizing calculated apparent KD values for His6-SUMO-She3p mutants. Binding affinities of She3p(331-405)
mutated in positions K340A, R341A, F344A, and Y345A appeared to be worse than for R342A. These were not quantified (not determined, n.d.) due to the lack of distinct band shifts. Of note,
only R342 makes intra-molecular contacts. Since R342A shows reduced RNA binding, also this stabilizing interaction seems important for RNA recognition. All EMSA were performed in three
independent experiments. SUPPLEMENTARY FIGURE 9 CONTROL EXPERIMENTS FOR _IN VIVO_ STUDIES AND COMPARISON OF LES. (A) Western blots with anti-Myc antibody showing the expression levels of
wild-type and mutant versions of She3p in corresponding yeast strains. Glucose-6-phosphate dehydrogenase (G6PD) was used as loading control. Only the triple mutant She3p(K340A, R341A, Y345A)
showed no expression. Western blots with anti-She2p antibody shows similar expression level of the wild-type strain and a strain expressing the mutant She2p(E172A, F176A). Phosphoglycerate
kinase 1 (Pgk1) was used as loading control. Western blots were performed in two independent experiments from different cell extracts. (B) Comparison of predicted secondary structures from
_ASH1_ LEs. Specifically recognized, essential bases in the _ASH1_-E3 LE are highlighted in red (C1779/ C1813), and blue (U1780). Conserved cytosines can also be found in E1, E2A and E2B.
Based on secondary-structure predictions at least E2A and E2B are likely to have both cytosines in similar distance on the opposing strand of their stems. However, the base-specific position
U1780 in E3 (blue base) is not conserved in the other elements (green bases), indicating that different subclasses of LEs might exist. Since the structural rearrangements observed in E3
upon binding (Fig. 2c) are impossible to predict with current bioinformatics tools, experimental approaches are required to understand the exact modes of binding for other LEs. SUPPLEMENTARY
INFORMATION SUPPLEMENTARY TEXT AND FIGURES Supplementary Figures 1–9, Supplementary Tables 1–5 and Supplementary Notes 1–2 (PDF 2755 kb) SUPPLEMENTARY DATA SET 1
Edelmann-Quanification_in_vivo_experiments-Table 2.xlsx (XLSX 18 kb) RIGHTS AND PERMISSIONS Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Edelmann, F., Schlundt, A., Heym, R.
_et al._ Molecular architecture and dynamics of _ASH1_ mRNA recognition by its mRNA-transport complex. _Nat Struct Mol Biol_ 24, 152–161 (2017). https://doi.org/10.1038/nsmb.3351 Download
citation * Received: 22 June 2016 * Accepted: 07 December 2016 * Published: 16 January 2017 * Issue Date: February 2017 * DOI: https://doi.org/10.1038/nsmb.3351 SHARE THIS ARTICLE Anyone you
share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the
Springer Nature SharedIt content-sharing initiative