- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
Wildlife and plant diseases can reduce biodiversity, disrupt ecosystem services and threaten human health. Emerging pathogens have displayed a variety of spatial spread patterns due to
differences in host ecology, including diffusive spread from an epicentre (West Nile virus), jump dispersal on a network (foot-and-mouth disease), or a combination of these (Sudden oak
death). White-nose syndrome is a highly pathogenic infectious disease of bats currently spreading across North America. Understanding how bat ecology influences this spread is crucial to
management of infected and vulnerable populations. Here we show that white-nose syndrome spread is not diffusive but rather mediated by patchily distributed habitat and large-scale gradients
in winter climate. Simulations predict rapid expansion and infection of most counties with caves in the contiguous United States by winter 2105–2106. Our findings show the unique pattern of
white-nose syndrome spread corresponds to ecological traits of the host and suggest hypotheses for transmission mechanisms acting at the local scale.
Wildlife and plant diseases can reduce biodiversity, disrupt ecosystem services and threaten human health1,2,3. White-nose syndrome (WNS), a highly pathogenic infectious disease caused by
the cryophilic fungus Geomyces destructans, has emerged in North American Vespertilionid bats, and is currently spreading4,5,6,7 throughout the Eastern United States and Canada. WNS was
first observed in 2006 in Schoharie County, NY, and within 2 years spread to 15 counties within a ∼230 km radius4. The speed of this spread, together with early reports of mass mortality in
infected caves, has generated concern about massive depletion of currently common bat species, attendant loss of economic services, and transmission to vulnerable populations of threatened
and endangered species5,8,9,10. Although WNS has now been reported in 20 states and up to ∼2,220 km from its North American origin, the ecological and environmental determinants of these
epidemiological dynamics are poorly understood.
Understanding the mechanisms by which WNS will expand is crucial to management of infected and vulnerable bat populations and the conservation of threatened and endangered bat species.
Particularly important is determining where and how fast WNS will spread. Historically, emerging pathogens have displayed a variety of spatial spread patterns due to differences in host
ecology, including diffusive spread from an epicentre (West Nile virus11), stratified diffusion (rabies virus12), jump dispersal on a network (foot-and-mouth disease virus13) or a
combination of these (Phytophthora ramorum14). Diffusive spread is expected for pathogens with highly mobile host species (for example, migratory passerines that transport West Nile virus),
while jump dispersal is expected for patchily distributed species with unequal time scales of within- and between-patch transmission (for example, livestock farms that mediated
foot-and-mouth disease). Either of these patterns is consistent with the natural history of bats: cave-dwelling bats exhibit long-distance aerial migration15 and high levels of seasonal
mixing16,17, but bats in temperate North America are also associated with patchily distributed geologic formations, particularly karst outcrops of the Appalachian range and Interior Plateaus
and Highlands18. How WNS should be expected to spread is, therefore, an open question.
To determine which of these two scenarios predominates in the spatial spread of WNS, we used maximum likelihood (ML) estimation to fit a series of models to data on the county-scale
infection history of this epidemic. We started with a simple diffusion model and added complexity to create spatially structured networks and then complex heterogeneous networks where
patches differed in attributes. We hypothesized that: geographic complexity gives rise to corridors that facilitate spread along the dominant cave-bearing geologic formations, spread is
accelerated by high concentrations of caves, and spread is retarded in warmer areas due to the shorter duration of cold temperatures through direct and indirect influences on bat condition
and fungal growth. We tested these hypotheses through statistical comparison of alternative models where the probability that county i does not become infected from previously infected
county j had the general form:
The function f describes the inverse of transmission intensity from j to i, numerically representing alternative hypotheses about the determinants of transmission. To further examine
possible scenarios, we fit additional models of moderate complexity, as well as an epidemiological model19 and a complex radiation model20. The radiation model used the same general form as
above, but the epidemiological model directly calculated . Simulations forward in time allowed us to compare spatial characteristics and predict future scenarios of spread under assumptions
of our best-supported model. We found the best fit model was of moderate complexity and included covariates regarding the number of potential hibernacula and length of winter. Simulations
forward in time based on this model suggest that spread to counties with caves will peak by 2015–2016 in the contiguous United States. While climate change may negatively impact spread to
uninfected counties according to interpretation of coefficients, applying the model to currently available projections of scenarios did not slow spread or further restrict the geographic
expansion. This study suggests that a variety of hibernating bat species are at risk and that populations across the contiguous United States eventually may be exposed.
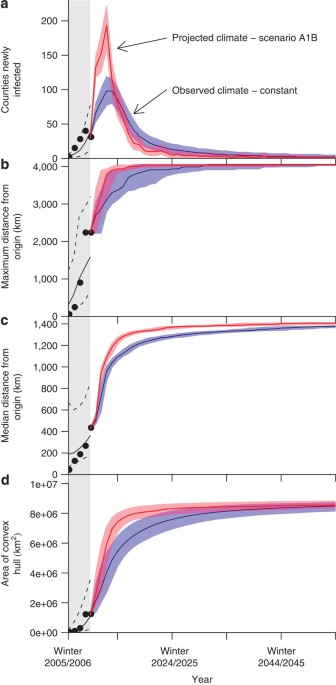
Inspection of summary statistics of observed data indicated that spread was variable over space and time in terms of four metrics (number of counties newly infected, maximum distance of
infected counties from the origin, median distance of infected counties from the origin, and area of the convex hull of infected counties, Supplementary Table S1). Particularly, the highest
number of new WNS infections and farthest spread occurred in the fourth epidemic winter and most newly infected counties (89%) have been relatively close (5 years, but reduced spread was
predicted into the Southeast and Southwest.
For spread continuing under constraints of these initial 5 years, the gravitycaves+winter model predicted the number of newly infected counties that contain caves will peak in the winter of
2013–2014 (95% prediction interval of 2013–2014 to 2014–2015). These projections suggest WNS will extend the maximum distance from the origin by 2032–2033 and will continue to fill in over
the next 73 years, such that 93% (±1%) of counties with caves will be infected by the winter of 2105–2106 (Supplementary Table S4). The predicted trajectory of spread is WNS will reach the
Rocky Mountains in the next 5 years before moving more slowly to the west coast (Fig. 3), whereas areas of high conservation concern in the Southeast and Texas are not predicted to become
infected until much later. Indeed, many of these counties and those in the Southwest went uninfected for >100 years in several simulations (Supplementary Fig. S16). Simulations using SU
parameter sets were consistent with these results (Supplementary Figs S31–S33, Supplementary Table S5). In addition, we considered possible effects of projected climate change by using
available simulations of future daily temperature. As North American winters have been relatively warm recently, these simulations predicted slightly faster spread of WNS despite the
expected increase in temperature overall (Fig. 1, Supplementary Figs S34, S35, Supplementary Table S6). However, the overall geographic characteristics of spread were consistent with the
observed winter data used to fit the model, despite differences in speed (Supplementary Figs S36–S41).
Mean estimated year of infection (yellow-to-red scale) under the gravitycaves+winter model for simulations using the ML parameter set begun from currently infected counties (2010–2011; blue
colour scale). Gray counties did not have reported caves or WNS observations. The model predicts spread to most (93%±1%) of the counties with caves in the contiguous United States within a
century of the first infection, many within the next 20 years. Spread to the southeast is slow as is spread to the southwest; longer time to infection in the Upper Midwest and New England
may be due to edge effects (Supplementary Information).
WNS has spread from a single New York county north to Maine, south to Alabama and west to Oklahoma. Simple diffusive models are not as well supported by data from the initial 5 years of WNS
spread as models that include spatial heterogeneity and a key environmental covariate, winter duration. Particularly, because a larger distance between two counties decreased the probability
of infection (that is, parameter β1 was positive for all fit models), the presence of geographic corridors due to the spatial pattern of counties with caves accelerated spread of WNS.
Further, in generalized gravity models, the parameter β2 was always positive indicating that the probability of infection increased with increasing number of caves in each county (Table 1).
This suggests that density-dependent transmission at higher bat population densities may facilitate spread by increasing contact rate, possibly during swarming. Furthermore, a county-level
climate variable, length of winter, was an important predictor and the related parameter, β3, was always negative indicating increased transmission probability to colder counties (Table 1).
This pattern may reflect the fungus’s physiological tolerances or increased bat survival in longer summers. This result is consistent with the conclusion that WNS mortality is predictable
under certain environmental conditions within Appalachia21.
There has been a great deal of interest in the effect of species diversity on spread of multi-host pathogens. Inclusion of species richness improved performance when added to the basic
gravitycaves model, and the positive coefficient might be interpreted to suggest that higher values of species richness retard the spread of WNS. However, this model is equally well
supported as the gravitycaves+northing model, possibly reflecting the latitudinal gradient of species richness along the eastern seaboard. Further, substituting the complete bat species
richness covariate with only those bats that hibernate did not improve fit to the observed data. Ultimately, species richness did not provide additional information when included with the
winter covariate in a gravity model. Future studies seeking to elucidate the role of species richness in G. destructans transmission should include information about the specific regions or
locations where hibernation occurs, that is, the spatial scales at which species ecological interactions may occur.
Thus, overall, our model suggests that the pattern of WNS spread results from a combination of both spatially diffusive mechanisms (that is, migratory events and movement between
hibernacula) and spread on a network (that is, clustering of habitats), and that there exist within- and between-region interactions that facilitate pathogen expansion. Empirical data
support this conclusion as small-scale features of habitat, species occurrence and species traits can mediate the appearance within hibernacula22. Our modelling framework cannot account
explicitly for these factors, but does not discount such factors as important mechanisms of spread.
Forward simulations initialized with an infection in Schoharie County, New York captured many features of spread observed in data, but were unable to correctly predict year of infection
within quartiles for some counties, consistent with other simulation studies23. Our goal in this study was to compare simple diffusive and geographically complex spread models that captured
the characteristics of WNS expansion; the observed pattern of occurrence is one potential realization of the spread dynamics and stochastic processes make predictions for individual counties
challenging and possibly ineffectual. Counties that were regularly under-predicted were those in Oklahoma and Missouri, which we consider to be possibly anomalous. Particularly, the
occurrence in Oklahoma represents a single live bat that tested positive for the pathogen and the pattern of infection differed from bats from the East Coast24. We also note that bats
readily use anthropogenic features25 that also can be suitable habitat for G. destructans, which would facilitate spread and thus bias our predictions where our proxy for available habitat
(cave density) is incomplete. However, recent analysis of WNS spread in the northeastern US suggests that mines are less likely to become infected than caves26, suggesting that this bias may
not be as great as one might fear. Thus, counties without caves but with mines may incur infection, but that the effect of these infections on spread will probably be small.
By simulating the effect of imperfect detection on model evaluation, we found that high levels of under-detection did not substantially bias summary statistics. Furthermore, it is likely
that detection following the first 2 years was very high given the profile of the disease. As we considered data at the county-level using a compartmental model, not the level of the
individual cave, we reduced the chance that we missed a critical data point while simultaneously removing potential issues of prevalence and within-county dynamics. Accordingly, we found our
results to be very robust to probable levels of under-detection.
It has been reported that the management of other emerging diseases would have benefited from earlier predictions of spread27. Our study sought to examine how the epidemic will progress
under the constraints and assumptions of the gravitycaves+winter model using ML and SU parameter sets by extrapolating from the state of infection as of 1 June 2011. These simulations
provide both general properties of anticipated spread (that is, approximate time frame for invasion of biologically sensitive areas), and specific estimates (with prediction intervals) for
all counties in the contiguous United States. Short-term predictions (that is, those within 10 years) are both most reliable and relevant to actions that might be taken immediately;
long-term predictions (that is, those well into the future) are less certain due to demographic stochasticity and potential population collapse9,10, as well as uncertainty about future
climate conditions28 and future control measures. Our choice in simulation duration (to 2105 under stable climate and 2100 for climate change scenarios) reflects the temporal extent of
available climate change forecasts, not necessarily considerations about a priori expectations of the progression of WNS epidemic. Additionally, because these predictions assume that
regionally different host communities do not differentially affect spread, future spread may be different than predicted in proportion to inhomogeneity in host communities and/or the extent
that resistance to infection is evolved22,29.
What will happen next? Our forward simulations from the observed distribution of WNS under ML parameters predicted that infection rate of new counties will peak in the winter of 2013–2014
(+1 winter) and that WNS will reach the maximum distance from Schoharie County in the winter of 2032–2033. Under the assumption that the first 5 years of spread are indicative of the future
pattern, our models predicted spread will continue rapidly along and within the Appalachians, filling in counties that were jumped over until reaching the southeast. As a result, potentially
susceptible species such as the Indiana Bat (Myotis sodalis) and Gray Bat (M. grisescens) will be of concern due to their already endangered status. Spread will be slow into the western
United States, although detection in Colorado may occur as soon as 2013–2014. This reduced spread in the projection is likely a trade-off between the number of caves and the length of winter
(Fig. 3). Species that have large geographic ranges, such as the Little Brown Bat (M. lucifigus), may see large population declines or extirpation at large scales9. Speculatively, species
richness across the contiguous United States may be reduced and community structure will shift in favour of non-cave hibernating bats30,31.
Predictions of spread using three climate change scenarios showed accelerated spread of WNS relative to projections based on observed data, with two caveats. First, the coarse resolution
reduces the climatic variability that is consistent with mountainous regions, which coincide with the distribution of caves. Thus the projected spread rates in some regions may be reduced or
accelerated by climate change in ways hard to capture with climate projections. Second, the projections had many more long winters than the observed data set, which could be due to
specifics of the climate model and year-to-year variability in climate projections (Supplementary Figs S42–44). As temperature appears to have a significant effect on WNS spread, actual
climate, especially the amount of cold weather in the immediate future, has the potential to significantly alter rate of spread and locations infected.
A potentially important puzzle piece missing from this analysis is spread within Canada. Absence of Canadian cave data (needed to estimate the cave density covariate) precluded incorporating
any such infection records into our models. We suspect including such information would have decreased the estimated time to infection in Northeast counties and in the Upper Midwest, but
would have had little impact on the spread into the Interior Plateaus and Highlands. It is possible that Canada could provide another route for long-distance dispersal events to reach the
West. However, Canadian winters may prove to act as a barrier to dispersal because of increased mortality of bats associated with longer winters, thus limiting the number of potential
dispersers.
Our model is the first of which we are aware to fit WNS occurrence data to broad geographic features and identify key characteristics of spread throughout the United States. An important
area for further development is construction of models representing the biological mechanisms that mediate dispersal and spread32. Unfortunately, key quantities such as bat-to-bat
transmission probability, recovery rate, frequency of recovery, lethality and between-cave mixing at hibernacula remain unknown to us. Recent experimental evidence7 shows that direct contact
can spread the fungus from infected to uninfected bats during hibernation and that airborne exposure within artificial hibernacula does not lead to infections in susceptible individuals.
Remarkably, without such a mechanistic understanding of spread, our best model matched all investigated aspects of the spatial dynamics of the epidemic, including maximum distance and number
of counties infected. Although diffusive spread may occur at small scales26, our results show that this spread process does not scale to larger spatial extents.
Our results also support an important mechanistic hypothesis: because of the relative importance of cave density in our models, we hypothesize that transmission is density-dependent at the
spatial scale of infection between caves. A cave-scale proportional hazard analysis found larger colonies were more likely to have become infected, but could not distinguish between the
effect of population density and the alternative that perhaps habitats preferred by bats are also more suitable for the fungus26. Lorch et al.7 speculate that swarming behaviour is a likely
mechanism for spread, whereby transmission between bats occurs during these fall periods of activity. As our model uses discrete time steps, it is consistent with a variety of transmission
modes including the swarming hypothesis. Lorch et al.7 provide evidence that direct exposure to conidia leads to infection within 102 days, but bat-to-bat experiments used infected bats
already presenting fungal growth. It is unclear, then, if autumn swarming is indeed the dominant mechanism of transmission, as other evidence shows infection clears during the summer33,34
and that the percentage of infected bats during autumn is very low7. We also can speculate on the effect of winter in the best-supported model. Winter length may increase transmission due to
increased conidia density on surviving bats or in caves. Alternatively, longer summers may slow spread, perhaps due to increased survivorship or reduced time for the syndrome to manifest as
found in infected individuals that have recovered in laboratory34 and field settings33. More mechanistic models of spread, such as Susceptible-Infected-Recovered models of transmission
among individual bats, could determine the importance of these effects5,32 and others, such as the postulated effect of cave microclimate (humidity) on bat infection and mortality22,26.
Unfortunately, data are currently too sparse to effectively estimate relevant parameters for transmission at the spatial extents considered here. Collection of individual incidence is,
therefore, a priority for future empirical work35.
Our model provides evidence for a non-diffusive spread pattern of WNS, where physical geography and environmental heterogeneity have a role in the speed and direction of spread. For other
recent notable pathogens, both introduced and emergent, host ecology influences the characteristics of spread, and the combination of ecological traits exhibited by bats matches the unique
mixture of heterogeneous and locally diffusive spread found here. Simulations forward suggest that spread will slow in the near future, but it will eventually reach most of the contiguous
United States. Bat diversity and community structure will be affected across all of North America if the spread along the Appalachians is representative of future spread. Vespertilionid bats
could be in critical danger if resistance, whether through an anthropogenic source or through evolution, does not soon emerge9,10,36.
Our model follows a compartmental strategy, similar to epidemiological Susceptible-Infected-Recovered models, where within-county prevalence and its potential role in spread to another
county are modelled implicitly. This framework permits hypothesis testing to statistically reject potential models of spread without making assumptions regarding mechanisms determining
within-county dynamics. Thus, our goal was to characterize broad geographic features that have a role in spread rather than to determine mechanisms that mediate local transmission.
Our simplest hypothesis of diffusive spread without geographic complexity was investigated by fitting a model where the probability of infection between all candidate sources j and
susceptible counties i declined with Euclidean distance between the centroids of i and j in an Albers equal-area projected data set of the contiguous United States through a logistic
function (Supplementary Equation S1). This model represents the belief that new infections are more probable in susceptible counties near infected counties. This model was modified to test
for distance-decay with additional long-distance dispersal by adding a fitted parameter as an exponent of the distance between counties (Supplementary Equation S8). We fit parameters of
these models using all counties in the contiguous United States, followed by fitting them to a restricted number of counties defined by the presence of caves (see above). If our proxy for
available hibernacula (that is, cave density) was irrelevant, then we would expect that model to fit the data equivalently or more poorly than the complete county data set. To test the
hypotheses that spread was accelerated by high concentrations of caves, we fit a model that included a generalized ‘gravity’ term38,39, where the connection between counties i and j was
weighted by the area of the county or density of caves in each (Supplementary Equations S15, S20). This model is a generalized version of the standard gravity model in that strength of the
connection was tuned by a fit parameter. This model was compared with diffusion on the restricted county network. We then tested for an effect of bat species richness by adding a term to the
gravitycave model (Supplementary Equation S21). Further, we tested for an effect of the number of hibernating bat species that may occur in a county during the year (Supplementary Equation
S22). To test the hypothesis that spread was retarded in warmer areas, we included winter duration (estimated number of days where minimum temperature was







