- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT To identify known and novel CYP4V2 mutations in patients with Bietti crystalline cornea (BCD), expand the spectrum of CYP4V2 mutations, and characterize the population history of
the c.802-8_810del17insGC mutation common in Asian populations, genomic DNA was isolated from peripheral blood samples from 58 unrelated patients with clinical diagnoses of BCD. Exons and
flanking intronic regions of the _CYP4V2_ gene were dideoxy DNA sequenced. Nonpathogenic polymorphisms were excluded and known mutations were identified by sequencing 192 unaffected
individuals from similar ethnic backgrounds and examination of online databases. The age of the c.802-8_810del17insGC mutation was estimated using three independent approaches. A total of 28
CYP4V2 mutations, 9 of which were novel, were detected in the 58 patients with BCD. These included 19 missense, 4 nonsense, 2 deletion, 2 splice site, and 1 insertion-deletion mutations.
Two missense variants of uncertain significance were also detected. The age of the c.802-8_810del17insGC mutation was estimated to be 1040–8200 generations in the Chinese and 300–1100
generations in the Japanese populations. These results expand the mutation spectrum of CYP4V2, and provide insight into the origin of the c.802-8_810del17insGC mutation in the Chinese
population and its transmission to the Japanese population. SIMILAR CONTENT BEING VIEWED BY OTHERS NATIONWIDE GENETIC ANALYSIS OF MORE THAN 600 FAMILIES WITH INHERITED EYE DISEASES IN
ARGENTINA Article Open access 22 May 2023 PHENOTYPIC DIVERSITY OBSERVED IN A CHINESE PATIENT COHORT WITH BIALLELIC VARIANTS IN BARDET-BIEDL SYNDROME GENES Article 08 April 2023 A
MULTI-ANCESTRY GWAS OF FUCHS CORNEAL DYSTROPHY HIGHLIGHTS THE CONTRIBUTIONS OF LAMININS, COLLAGEN, AND ENDOTHELIAL CELL REGULATION Article Open access 06 April 2024 INTRODUCTION Bietti
crystalline cornea-retinal dystrophy (BCD, MIM210370) is a progressive autosomal recessive retinal dystrophy first reported by Bietti in 1937. It is characterized by numerous small
glittering yellow-white crystals at the posterior pole of the retina associated with progressive atrophy of the retinal pigment epithelium (RPE), pigment clumps, and choroidal sclerosis.
Some patients have similar crystal deposits at the corneoscleral limbus and in circulating lymphocytes and skin fibroblasts.1 Most patients begin to show night blindness, decreased visual
acuity, and paracentral scotomas between the second and fourth decades progressing to peripheral visual field loss and marked visual impairment.1 BCD is more common in East Asia, especially
in China and Japan.2 Patients show abnormally high triglycerides and cholesterol storage in cultured cells from BCD patients, lack two fatty acid-binding protein activities,3 and have
decreased metabolism of labeled fatty acid precursors into _n_-3 polyunsaturated fatty acids (_n_-3PUFA).4 Sequence variants that affect the function of CYP4V2 (MIM 608614), a member of the
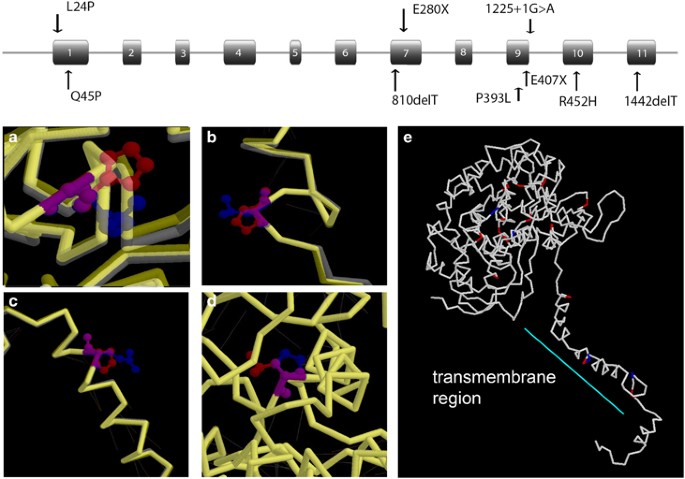
heme thiolate cytochrome P450 subfamily 4 (CYP4), predominantly active in fatty acid metabolism, cause BCD.5 The _CYP4V2_ gene spans 21 kb, comprising 11 exons encoding a 525 amino acid
protein (Figure 1, top). Although CYP4V2 is expressed in almost all tissues, it is expressed at high levels in the retina and RPE, and at somewhat lower levels in the cornea tissues, which
show the major clinical findings of BCD.5 CYP4V2 is a PUFA hydroxylase highly expressed P450 in the transformed human RPE cell line ARPE-19.6 Consistent with this, abnormalities in ω3-PUFAs
and their metabolism have been demonstrated in patients with BCD.4, 7, 8 The retinal findings and systemic lipid abnormalities have been recapitulated in a knockout mouse model.9 Before this
study, 73 mutations in CYP4V2 have been described in the literature and clinical databases. To expand the spectrum of these CYP4V2 mutations in patients with BCD, the _CYP4V2_ gene was
sequenced in 58 patients diagnosed with BCD and their sequences were compared with those of 192 unrelated healthy controls as well as known mutations and sequence changes in online sequence
variation databases, identifying 28 different mutations (9 novel) and 2 sequence variants of unknown significance, and delineating the origin of the common c.802-8_810del17insGC indel
mutation. MATERIALS AND METHODS PATIENTS This study was approved by the CNS Institutional Review Board of the National Institutes of Health and consent obtained in accordance with the
Declaration of Helsinki. Patients were diagnosed with BCD on the basis of clinical features,1 and were of European, Chinese, Palestinian Arabic, and Korean ancestry (Table 1). Control
individuals were of matched ethnic backgrounds. DNA AMPLIFICATION AND MUTATION DETECTION Genomic DNA was extracted directly from blood or transformed lymphoblastoid cell lines by standard
phenol–chloroform protocols.10 PCR amplification of _CYP4V2_ exons 1–11 including intron–exon boundaries and 50 bp of flanking sequence was performed as reported previously.5 PCR products
were purified using Agencourt CleanSEQ (Beckman Coulter, Biomek NX, Brea, CA, USA). Sequencing was performed on an ABI PRISM 3130 Automated sequencer (Applied Biosystems, Foster City, CA,
USA) and analyzed using Mutation Surveyor v3.30 (Soft Genetics, State College, PA, USA) or Lasergene 8.0, (DNASTAR, Madison, WI, USA). Control DNA samples from 192 unrelated individuals of
Chinese, Japanese, or European ethnic origin were also analyzed and the 1000 Genomes Database (http://www.1000genomes.org/), NCBI dbSNP database (http://www.ncbi.nlm.nih.gov/projects/SNP/),
NHLBI Exome Variant Server (http://evs.gs.washington.edu/EVS/), Biobase (https://portal.biobase-international.com) and ExAC Database (http://exac.broadinstitute.org/) were also searched for
all novel sequence variations. Effects of potential splice mutations were modeled using online resources including neural networking through The Berkeley Drosophila Genome Project
(http://www.fruitfly.org/seq_tools/splice.html) and the Center for Biological Sequence Analysis (http://www.cbs.dtu.dk/services/NetGene2/), consensus values at the Inserm Human Splicing
Finder (http://umd.be/HSF/), the Alternative Splice Site Predictor (ASSP, http://wangcomputing.com/assp/index.html), and MaxEntScan
(http://genes.mit.edu/burgelab/maxent/Xmaxentscan_scoreseq.html). Mutations are described with reference to the CYP4V2 mRNA sequence NM_207352.3 and gene sequence NG_007965.1. Chromosomal
positions are based on hg19/GRCh37. Variants were submitted to the LOVD (http://databases.lovd.nl/shared/genes/CYP4V2) with screening (submission) numbers 0000081248- 0000081331. MOLECULAR
MODELING Modeling of the novel CYP4V2 mutant proteins was performed by homology modeling based on predicted coordinates for the wild-type CYP4V2 protein5 using the SWISSMODEL Workspace
web-based environment with default values,11, 12, 13 and visualized using RASMOL.14 HAPLOTYPE ANALYSIS AND AGE ESTIMATION Delineation of SNP haplotypes in individuals carrying the
c.802-8_810del17insGC mutation was done by examination as most individuals were homozygous. The haplotype was extended until half of the individuals in the Chinese or Japanese populations
diverged. Haplotypes were grouped by country and were sorted into those requiring one or two recombination events in their descent from the ancestral haplotype by examination to minimize the
number of recombination events required. The mutation age was estimated as described in Equation (1) of Risch _et al_,15, 16 , where _δ_ is the linkage disequilibrium constant,
_δ_=(_P_D−_P_N)/(1−_P_N), with _P_D being the frequency of the allele on chromosomes of mutation carriers and _P_N being the frequency of the allele in the control population. Genotypes of
unaffected individuals were taken from the 1000 Genomes database (103 Han Chinese in Beijing: CHB and 105 Southern Han Chinese: CHS for the Chinese population; 104 Japanese in Tokyo: JPT for
the Japanese population). Haplotypes were estimated using the CHM method as implemented in the Golden Helix SVS (Golden Helix, Bozeman, MT, USA). The map distances were inferred on the
basis of the physical distances as given in GRCh37/hg19 from the UCSC Genome Browser assuming 1 Mb=1 cM. Alleles at SNPs showing no recombination in cases also were collapsed into a single
haplotype and analyzed as a single marker in the two marker approach described in Equation (2) of Risch _et al._ In addition, marker specific values of g were estimated using the method
described in Equation 1 of Goldstein _et al._17 _K_=_cR_+μM+(1-_c_-_μ_)_I_, where _c_ and _μ_ are the recombination and mutation rates, respectively, _R_ is a 2 × 2 matrix with R11=R12=_a_,
and R21=R22=1−_a_, where _a_ is the frequency of the ancestral allele in the control population, _M_ is a 2 × 2 matrix with M11=0, M12=1/3, M21=1, and M22=2/3, to account for the frequency
with which a mutation might remove an ancestral allele (all cases) or move to an ancestral allele (1 of 3 possible bases), and _I_ is the 2 × 2 identity matrix. The original frequency vector
is (1, 0) as it occurs on the founder haplotype, and this association is reduced by multiplying by _K_ at each generation (_g_) until the current frequency of the ancestral allele is
reached. Finally, the marginal posterior probability distribution of the age18 of the c.802-8_810del17insGC was estimated using the DMLE+2.2 software developed by Reeve and Rannala.19 This
program estimates the age in generations by comparing the observed haplotypes in chromosomes from affected and unaffected sample sets considering the map distances, the population growth
rate (gen_r_), and the proportion of the mutation bearing chromosomes sampled, but has a strong dependence on assumptions regarding the population history of China and Japan. Somewhat
arbitrarily, population growth was modeled since 2300 BC, the earliest time at which reliable figures were available for both populations, although not as old as estimates of the mutation
age, assuming 25 year generations. RESULTS MUTATION SCREENING OF THE _CYP4V2_ GENE Mutation screening of CYP4V2 was performed in 58 patients with BCD. A total of 28 mutations and 2 variants
of uncertain significance were identified (Table 1 and Figure 1). Of the nine novel mutations, five were in homozygous and 4 in compound heterozygous cases (Table 1A). Patients 6295 and 6296
were twins; the other 10 cases were sporadic affected individuals. Clinical details of cases with novel mutations are given in Supplementary Table S1. Structural predictions of the effects
of the four novel missense mutations were analyzed by homology based on coordinates for the wild-type CYP4V2 protein (Figure 1, bottom). * 1) c.71T>C, p.(Leu24Pro) produces minimal change
in the backbone of the protein, and no hydrogen bonds are disrupted. However, the potential of this variant to affect _CYP4V2_ function and its high conservation across species possibly
relates to its position in the transmembrane region, which would be highly sensitive to amino acid changes, and especially a change to a proline, which would disrupt the _α_-helical
structure of this region (Figures 1b and e). Leu24 is conserved among seven of the aligned species, although absent from the horse and zebrafish (Table 2), and p.(Leu24Pro) has blosum 62 and
Grantham scores of −3 and 98, respectively.20, 21 * 2) c.134A>C, p.(Gln45Pro) Gln45 is not completely conserved in all nine species (Table 2), although the substitutions are conservative
in all species except the chicken. It has a blosum 62 and Grantham scores of −1 and 76, respectively. As with the c.71T>C, p.(L24P) mutation, the disagreement in the predictive programs
and high conservation of this residue might relate to its position in the transmembrane segment, where it distorts the _α_-helix, slightly disrupting two hydrogen bonds (Figures 1c and h). *
3) c.810delT, p.(Glu271Argfs*34) results in a premature stop in an internal exon, so that the mRNA is predicted to undergo nonsense-mediated decay (NMD). * 4) c.838G>T (GAA>TAA),
p.(Glu280*) also should result in NMD. * 5) Although c.1178C>T, p.(Pro393Leu) produced little distortion of the protein backbone in this region and disrupted no hydrogen bonds (Figure
1d), it predicted to be damaging because of its position near the porphyrin ring and heme group of the active site of the protein. Pro393 is highly conserved and has blosum 62 and Grantham
scores of −3 and 98, respectively. * 6) c.1219G>T, p.(Glu407*) should result in NMD. * 7) c.1225+1 G>A, p.(Gly364_V408del) is predicted to skip exon 9. * 8) c.1355G>A, p.(Arg452His)
is predicted to interfere with coordination of the heme ring and binding of the fatty acid substrate (Figures 1a and e). Arg452 is conserved in all species examined from human to zebrafish
(Table 2). * 9) c.1441delT, p.(Ser481Argfs*4) would not result in nonsense-mediated decay, but would substitute 4 random amino acids for the last 44, destabilizing the active site. The
remaining 28 mutations, including 19 (68%) missense, 4 (14%) nonsense, 2 (7%) splice, 2 (7%) deletions, and a single indel (4%), have been reported previously (Table 1B). Although the
nonsense, splice, deletion, and indel mutations would be predicted to have a severe effect wherever they occurred, the missense mutations might be expected to identify parts of the protein
structure particularly susceptible to perturbation. As can be seen in Figure 1e, the missense mutations taken as a whole cluster in two regions: the transmembrane region, without which the
CYP4V2 protein would not be expected to insert into the membrane, and the area around the porphyrin ring, including residues that might influence coordination of the heme group and those in
the _α_-helices that support and position them. ETHNIC ORIGINS OF CYP4V2 MUTATIONS IN BCD PATIENTS Although the number of mutations identified in this study and Li _et al_,5 40, is
relatively small, some patterns do emerge when the ethnicity of the patients displaying those mutations is examined, especially the more common mutations. Of the 99 combined unrelated
patients and families examined, the ethnicity of 13 of them was unknown, leaving 86: 31 of European origin, 38 of Chinese origin, 1 of Arabic origin, 8 of Korean origin, and 8 of Japanese
origin (Table 3). Although there is some overlap, specific mutations are generally restricted to Asian (Chinese, Korean, and Japanese) or European populations. Of these, 22 patients showed
the c.802-8_810del17insGC indel mutation distributed among the Chinese, Korean, and Japanese patients. This mutation was not seen in European or Arabic populations. The next two most common
mutations (seen in 10 patients each) were the c.992A>C mutation, seen in Chinese and Korean populations, and the c.1091-2A>G mutation, restricted to Chinese patients. In contrast, the
c.332T>C mutation, seen in 8 patients and the c.64C>G mutation were restricted to 3 Europeans. The c.367A>G, c.694C>T, c.1199G>A, and c.1328G>A mutations were seen in both
European and Chinese populations, whereas the remaining mutations, although reported previously, occurred only a single time in this study. HAPLOTYPE ANALYSIS AND HISTORY OF THE
C.802-8_810DEL17INSGC INDEL MUTATION Intragenic SNP haplotypes from the Chinese and Japanese BCD patients harboring the c.802-8_810del17insGC indel mutation included in this study were
aligned along with those from Li _et al_,5 Lin _et al_,22 and Lee _et al_23 as shown in Figure 2. The CAAT(delCT)TA(indel)TCA haplotype shaded in dark blue in homozygotes was the most common
in Chinese, Japanese, and Koreans, and was assumed to be the ancestral haplotype for all three population groups.22 Individual alleles of the SNPs composing the founder haplotype were
highly conserved among patients: rs7663027 (74% C), rs10013653 (100% A), rs7682918 (94% A), rs12507156 (94%T), rs397722245 (83% deletion), rs4862662 (94% T), rs13146272 (94% A), rs207482233
(100% del—the c.802-8_810del17insGC indel), rs28698123 (100%T), rs7667777 (68% C), and rs2276918 (51% A), although some lie beyond divergent alleles closer to the mutation and thus are
identical by state rather than descent. The entire founder haplotype was found in 0 and 30% of Chinese controls and patients and 0 and 21% of Japanese controls and patients, respectively,
for a _P_=0.0039 and 0.00676 in Chinese and Japanese populations. In fact, none of the partial founder haplotypes shown in Figure 3 were estimated to be present in the Japanese population,
whereas only the partial founder haplotype CAAT(CT)TATCG, was estimated to occur in 4% of the Chinese control population. Because of the availability of pedigree information and the
assumptions made in Haplotyping the families, the EM-estimated frequencies of the founder haplotype differ from those seen in our study (44 and 29% in Chinese and Japanese affected,
respectively). In Figure 3a the BCD-associated haplotypes from all three populations are ordered into three levels based on the number of recombination events required to derive each
haplotype from the founder, which is shown at the top (level 0). A single recombination event can generate the three haplotypes shown in level 1 (one above and two different events below the
mutation). From these three haplotypes the remainder, seen on level 2, can be generated by a second recombination on the opposite side of the mutation from the first. An alternative,
although more complex, pathway to the two haplotypes on the left side of level 2 from the two haplotypes on the right of level 1 is shown with gray arrows. Thus, all haplotypes bearing the
c.802-8_810del17insGC mutation can be generated from the founder haplotype by a maximum of two recombination events, although additional recombination events occurring beyond those shown in
Figure 3a would not be identifiable from this analysis, and mutation of a SNP, while infrequent, could not be differentiated from a recombination event. In Figure 3b the haplotypes are
arranged in order of their divergence from the founder haplotype with the number of individuals (or families) and the population to which they belong shown above each haplotype. The founder
haplotype is the most common in the Chinese population, whereas the founder haplotype and a second haplotype with a single C>G change are both common in the Japanese population. Finally,
a haplotype differing from the founder by having G alleles for the final two SNPs, rs7667777 and rs2276918, accounts for all c.802-8_810del17insGC mutations seen in the Korean population.
This haplotype is also seen in a single family in both the Japanese and Chinese populations. The preservation of the founder haplotype in all three populations strongly suggests a single
common origin for the c.802-8_810del17insGC mutation, as has been suggested by Lin _et al._22 In addition, the greater diversity (12 obligate recombination events in the Chinese _vs_ 6 in
the Japanese haplotypes) and occurrence of recombination events closer to the mutation itself suggest that the mutation might have existed in the Chinese population longer than it has in the
Japanese population. Although the available sample set does not include enough individuals of Korean origin to provide strong support, the existence of a single haplotype would suggest that
the c.802-8_810del17insGC mutation might have been introduced into this population relatively recently. As this haplotype is present in both Chinese and Japanese populations, it is not
possible to determine from which it originates. To examine the origin of the c.802-8_810del17insGC mutation more closely, the mutation age was estimated in the Chinese and Japanese
populations using the approach described by Risch _et al_,15, 16 and Goldstein _et al_,17 as well as Bayesian disequilibrium estimates of the marginal posterior density as instituted in the
DMLE+ program.19 A summary of these results with the age given in generations is shown in Table 4. Using the method described by Risch _et al_ for single markers, estimates ranged from 1150
to 7044 generations, whereas the multiple marker approach using markers combined into haplotypes gave estimates of 3400 and 4500 generations with left and right flanking markers,
respectively. Using the method described by Goldstein _et al_, a similar range was obtained for the single markers, 1841–8200, and 1900 for the rs10013653-rs12507156-rs12507156-rs397722245
haplotype. With both methods, the estimates obtained with rs7663027 is significantly higher than that obtained with the other markers. One possible explanation for this is that the founder
allele frequency at this marker, 0.5625, is beginning to approach the allele frequency in the control population, 0.475, suggesting that the distance between this marker and the mutation
might be on the upper limits for usefulness in this estimation. Finally, the estimate using DMLE2 assuming population growth rates from 2300 BC, the earliest time reliable estimates are
available for this population, is 1040 with 95% confidence limits of 931–1237. When the age of the mutation in the Japanese population was estimated, values of 708 and 933 generations were
obtained with rs7667777 and rs2276918, respectively. A multi-marker analysis of the rs28698123-rs7667777 and rs28698123-rs7667777-rs2276918 haplotypes yielded an estimate of 1100
generations, and the method described by Goldstein _et al_ provided estimates of 933 and 300 generations for rs7667777 and rs2276918, respectively. The estimate from DMLE2 using population
growth from 2300 BC was 624 generations with 95% confidence intervals of 538–738 generations. In the Japanese population, estimates could not be made with many markers because there was no
recombination within the haplotype or, for rs7663027 the founder allele frequency was actually below that in the general population. The population age in the Korean population could not be
calculated because of the small number of risk haplotypes available and the absence of available control haplotype data. DISCUSSION In the current study, we report the results of analysis of
58 unrelated patients with BCD, identifying 9 novel mutations (Table 1), a significant addition to the 82 mutations currently listed in HGMD. One of the limitations of this study is the
paucity of precise clinical data sufficient to completely characterize some of the patients. However, the available data suggest that all the patients have typical BCD, which is perhaps
related to their being referred specifically for sequencing of the _CYP4V2_ gene. Similarly, from the clinical data available it was not possible to correlate any specific genotype with a
phenotypic characteristic or severity. This is consistent with previous studies showing a high degree of clinical variability within and between families, without an obvious correlation to a
specific mutation, and also is not surprising, as each of the identified mutations is predicted to result in complete absence of function in CYP4V2. Overall, 19 missense (68%), 1 indel
(4%), 4 nonsense (14%), 2 deletions (7%), and 2 splice site mutations (7%) were identified in this study (28 mutations in all). These results are very similar to mutation frequencies
tabulated in HGMD (missense and nonsense, 68% indels, 1% deletions, 12% insertions 3% gross deletions 2% and splice site mutations, 14%). The CYP4V2 protein structure, as estimated by
homology modeling, is predicted to have a transmembrane segment at the amino terminal end, followed by a link to the globular domain, consisting of 18 _α_-helices connected by _β_-sheet and
random coil structures.5 Although the splice, frameshift, and nonsense mutations are predicted to cause gross structural deformities of the protein fold, the missense mutations are seen to
cluster in two areas. One is the transmembrane segment, which should interfere with insertion of the enzyme into the membrane. The other is the active site, in which mutations would disrupt
coordination of the porphyrin ring required for enzymatic activity or an _α_-helix supporting this ring. The c.1355G>A (p. Arg452His) mutation is listed as novel, because although it is
present in heterozygous form in the ExAC database (mostly in Europeans), it has not previously been associated with BCD or described in any BCD patients. The previously described missense
variants c.64C>G, p.(L22V), and c.775A>C, p(Lys259Gln) were both found in both homozygous and heterozygous individuals (Table 1B). They are predicted to be tolerated by SIFT and benign
by PolyPhen2. They also have frequencies of 27–52% and 9–25%, respectively, in various populations on the 1000 Genomes database, so that they seem unlikely to affect CYP4V2 function. This
has been noted in previous publications although they do turn up frequently in BCD cases, and the c.775A>C, p(Lys259Gln) variant has been associated with deep vein thrombosis.22, 24, 25
Estimating the likelihood of novel missense variants to affect function is certainly less certain than null alleles, especially where there is disagreement between the predictive programs.
In part, this disagreement results from the different predictive algorithms used by each program. Polyphen-2 uses specific sequence (eg, active site or known motif), phylogenetic (species
alignment with position-specific independent counts or PSIC), and structural information to estimate the effect on the protein structure and function. PROVEAN predictions are based on
conservation of closely related sequences, whereas Condel combines the results of SIFT, PolyPhen2, MutationAssessor and FatHMM to predict the effects of an amino acid change. Thus, each of
these programs uses a slightly different combination of factors normalized to different databases, with varying degrees of sensitivity and specificity, so that some inconsistencies are to be
expected. However, their predictive accuracy certainly increases when all agree. Although the identified changes seem likely to be causative, it is possible that mutations in regulatory
regions outside the coding sequence cause the disease. We have tried to minimize this problem by sequencing the core promoter region (~100 bp) of CYP4V2 in all patients with novel missense
mutations, and no changes were identified (data not shown). Although it is theoretically possible that mutations in a different gene can cause BCD in these cases, no other gene or linkage
region has been suggested in multiple previous studies, many of which include linkage data, and two mutations have been found in ~92% of BCD patients.5, 22, 23, 24, 26, 27, 28, 29, 30, 31,
32, 33, 34 The c.802-8_810del17insGC change, the most common mutation of CYP4V2 in East Asian BCD patients, was found in 15 cases of 58 BCD patients in this study, all of East Asian origin.
Overall, we have found the c.802-8_810del17insGC change in 14 individuals of Chinese, 1 of Japanese and 4 of Korean origin,5 consistent with estimates from the 1000 Genomes database, in
which the allele frequency of this mutation is 0.01 in Japan and 0.005 in China, but only 0.002 in Europe and was not seen in African, South Asian, or American populations. In this regard,
BCD overall has been reported to be more common in East Asia populations with an estimated gene frequency of 0.005 in China.35 Estimation of the population age, origin, and population
history of found the c.802-8_810del17insGC mutation were based on our results and those of previous studies in which the risk haplotype was determined in Chinese and Japanese Bietti cases.5,
22, 23 The age of the mutation was estimated using three approaches, an analytical approach,15 an iterative approach,17 and a posterior Bayesian distribution approach as implemented in the
program DMLE+2.2.36 On the basis of their analysis of the intragenic SNP haplotypes, Lin _et al_ previously suggested that the c.802-8_810del17insGC had a common origin in the Japanese and
Chinese populations. Our analysis is completely consistent with that suggestion, especially as the ancestral risk haplotype was not identified in either Chinese or Japanese control
individuals, suggesting that it is quite rare in both populations. Although the age estimates vary widely, two findings are apparent: (1) the mutation is extremely old, occurring millennia
ago, and (2) the temporal precedence of the c.802-8_810del17insGC mutation in the Chinese population further suggests that the mutation originated in ancient China and after some time,
perhaps tens of thousands of years, was introduced into the Japanese population where it eventually attained the current allele frequency. It is difficult to know whether the Chinese or
Japanese population served as the source of this mutation in the Korean population, as there have been interchanges with both in recent and ancient history. The identical haplotype found in
Korea is found in a single instance in both the Chinese and Japanese patients, and we have only a limited number of Korean risk and no control haplotypes on which to base an age estimate.
However, these results must be interpreted with some caution, as they are sensitive to the parameters used in the estimation. Although physical distance between the SNPs is accurately known,
the associated recombination frequency was estimated from an average value for the genome. This probably would have minimal effects on the relative age of the mutation in the Japanese and
Chinese populations, as the same values were used for both estimates. Similarly, negative selection would probably not be a major problem in this recessive disease, especially as most BCD
patients maintain functional vision through the fourth or fifth decade, well beyond reproductive age.23, 26 Estimates of haplotype and genotype frequencies of the risk population should also
be robust, and the risk haplotypes include all those studied and published so far. The results are also consistent with the greater diversity of the risk haplotypes and presence of
recombination events closer to the mutation in the Chinese population. In conclusion, we report 28 CYP4V2 mutations, 9 of which were novel. These expand the spectrum of CYP4V22 mutations
associated with BCD, assisting with molecular diagnosis and providing additional insight into the structural importance of the variant amino acids. Although there is some overlap, most
mutations are confined either to the European or Asian population groups. In addition, these data suggest a single origin of the common c.802-8_810del17insGC mutation in the Chinese
population, followed by spread to Japan and then Korea. REFERENCES * Kaiser-Kupfer MI, Chan CC, Markello TC _et al_: Clinical biochemical and pathologic correlations in Bietti’s crystalline
dystrophy. _Am J Ophthalmol_ 1994; 118: 569–582. Article CAS Google Scholar * Hu DN : Prevalence and mode of inheritance of major genetic eye diseases in China. _J Med Genet_ 1987; 24:
584–588. Article CAS Google Scholar * Lee J, Jiao X, Hejtmancik JF, Kaiser-Kupfer M, Chader GJ : Identification, isolation, and characterization of a 32-kDa fatty acid-binding protein
missing from lymphocytes in humans with Bietti crystalline dystrophy (BCD). _Mol Genet Metab_ 1998; 65: 143–154. Article CAS Google Scholar * Lee J, Jiao X, Hejtmancik JF _et al_: The
metabolism of fatty acids in human Bietti crystalline dystrophy. _Invest Ophthalmol Vis Sci_ 2001; 42: 1707–1714. CAS PubMed Google Scholar * Li A, Jiao X, Munier FL _et al_: Bietti
crystalline corneoretinal dystrophy is caused by mutations in the novel gene CYP4V2. _Am J Hum Genet_ 2004; 74: 817–826. Article CAS Google Scholar * Nakano M, Kelly EJ, Rettie AE :
Expression and characterization of CYP4V2 as a fatty acid {omega}-hydroxylase. _Drug Metab Dispos_ 2009; 37: 2119–2122. Article CAS Google Scholar * Nakano M, Kelly EJ, Wiek C, Hanenberg
H, Rettie AE : CYP4V2 in Bietti’s crystalline dystrophy: ocular localization, metabolism of omega-3-polyunsaturated fatty acids, and functional deficit of the p.H331P variant. _Mol
Pharmacol_ 2012; 82: 679–686. Article CAS Google Scholar * Lai T, Chu KO, Chan KP _et al_: Alterations in serum fatty acid concentrations and desaturase activities in Bietti crystalline
dystrophy unaffected by CYP4V2 genotypes. _Invest Ophthalmol Vis Sci_ 2010; 51: 1092–1097. Article Google Scholar * Lockhart CM, Nakano M, Rettie AE, Kelly EJ : Generation and
characterization of a murine model of Bietti crystalline dystrophy. _Invest Ophthalmol Vis Sci_ 2014; 55: 5572–5581. Article CAS Google Scholar * Smith RJH, Holcomb JD, Daiger SP _et al_:
Exclusion of Usher syndrome gene from much of chromosome 4. _Cytogenet Cell Genet_ 1989; 50: 102–106. Article CAS Google Scholar * Bordoli L, Kiefer F, Arnold K, Benkert P, Battey J,
Schwede T : Protein structure homology modeling using SWISS-MODEL workspace. _Nat Protoc_ 2009; 4: 1–13. Article CAS Google Scholar * Arnold K, Bordoli L, Kopp J, Schwede T : The
SWISS-MODEL workspace: a web-based environment for protein structure homology modelling. _Bioinformatics_ 2006; 22: 195–201. Article CAS Google Scholar * Biasini M, Bienert S, Waterhouse
A _et al_: SWISS-MODEL: modelling protein tertiary and quaternary structure using evolutionary information. _Nucleic Acids Res_ 2014; 42: W252–W258. Article CAS Google Scholar * Sayle RA,
Milner-White EJ : RASMOL: biomolecular graphics for all. _Trends Biochem Sci_ 1995; 20: 374. Article CAS Google Scholar * Risch N, de Leon D, Ozelius L _et al_: Genetic analysis of
idiopathic torsion dystonia in Ashkenazi Jews and their recent descent from a small founder population. _Nat Genet_ 1995; 9: 152–159. Article CAS Google Scholar * Colombo R : Age and
origin of the PRNP E200K mutation causing familial Creutzfeldt–Jacob disease in Libyan Jews. _Am J Hum Genet_ 2000; 67: 528–531. Article CAS Google Scholar * Goldstein DB, Reich DE,
Bradman N, Usher S, Seligsohn U, Peretz H : Age estimates of two common mutations causing factor XI deficiency: recent genetic drift is not necessary for elevated disease incidence among
Ashkenazi Jews. _Am J Hum Genet_ 1999; 64: 1071–1075. Article CAS Google Scholar * Slatkin M, Rannala B : Estimating allele age. _Annu Rev Genomics Hum Genet_ 2000; 1: 225–249. Article
CAS Google Scholar * Reeve JP, Rannala B : DMLE+: Bayesian linkage disequilibrium gene mapping. _Bioinformatics_ 2002; 18: 894–895. Article CAS Google Scholar * Henikoff S, Henikoff JG
: Amino acid substitution matrices from protein blocks. _Proc Natl Acad Sci USA_ 1992; 89: 10915–10919. Article CAS Google Scholar * Grantham R : Amino acid difference formula to help
explain protein evolution. _Science_ 1974; 185: 862–864. Article CAS Google Scholar * Lin J, Nishiguchi KM, Nakamura M, Dryja TP, Berson EL, Miyake Y : Recessive mutations in the CYP4V2
gene in East Asian and Middle Eastern patients with Bietti crystalline corneoretinal dystrophy. _J Med Genet_ 2005; 42: e38. Article CAS Google Scholar * Lee KY, Koh AH, Aung T _et al_:
Characterization of Bietti crystalline dystrophy patients with CYP4V2 mutations. _Invest Ophthalmol Vis Sci_ 2005; 46: 3812–3816. Article Google Scholar * Shan M, Dong B, Zhao X _et al_:
Novel mutations in the CYP4V2 gene associated with Bietti crystalline corneoretinal dystrophy. _Mol Vis_ 2005; 11: 738–743. CAS PubMed Google Scholar * Bezemer ID, Bare LA, Doggen CJ _et
al_: Gene variants associated with deep vein thrombosis. _JAMA_ 2008; 299: 1306–1314. Article CAS Google Scholar * Meng XH, Guo H, Xu HW _et al_: Identification of novel CYP4V2 gene
mutations in 92 Chinese families with Bietti’s crystalline corneoretinal dystrophy. _Mol Vis_ 2014; 20: 1806–1814. PubMed PubMed Central Google Scholar * Yin H, Jin C, Fang X _et al_:
Molecular analysis and phenotypic study in 14 Chinese families with Bietti crystalline dystrophy. _PLoS One_ 2014; 9: e94960. Article Google Scholar * Lai TY, Ng TK, Tam PO _et al_:
Genotype phenotype analysis of Bietti’s crystalline dystrophy in patients with CYP4V2 mutations. _Invest Ophthalmol Vis Sci_ 2007; 48: 5212–5220. Article Google Scholar * Tian R, Wang SR,
Wang J, Chen YX : Novel CYP4V2 mutations associated with Bietti crystalline corneoretinal dystrophy in Chinese patients. _Int J Ophthalmol_ 2015; 8: 465–469. PubMed PubMed Central Google
Scholar * Xiao X, Mai G, Li S, Guo X, Zhang Q : Identification of CYP4V2 mutation in 21 families and overview of mutation spectrum in Bietti crystalline corneoretinal dystrophy. _Biochem
Biophys Res Commun_ 2011; 409: 181–186. Article CAS Google Scholar * Hayakawa M, Fujiki K, Kanai A _et al_: Multicenter genetic study of retinitis pigmentosa in Japan: I. Genetic
heterogeneity in typical retinitis pigmentosa. _Jpn J Ophthalmol_ 1997; 41: 1–6. Article CAS Google Scholar * Wada Y, Itabashi T, Sato H, Kawamura M, Tada A, Tamai M : Screening for
mutations in CYP4V2 gene in Japanese patients with Bietti’s crystalline corneoretinal dystrophy. _Am J Ophthalmol_ 2005; 139: 894–899. Article CAS Google Scholar * Yokoi Y, Sato K, Aoyagi
H, Takahashi Y, Yamagami M, Nakazawa M : A novel compound heterozygous mutation in the CYP4V2 gene in a Japanese patient with Bietti’s crystalline corneoretinal dystrophy. _Case Rep
Ophthalmol_ 2011; 2: 296–301. Article Google Scholar * Jin ZB, Ito S, Saito Y, Inoue Y, Yanagi Y, Nao-i N : Clinical and molecular findings in three Japanese patients with crystalline
retinopathy. _Jpn J Ophthalmol_ 2006; 50: 426–431. Article CAS Google Scholar * Hu DN : Ophthalmic genetics in China. _Ophthalmic Paediatr Genet_ 1983; 2: 39–45. Article Google Scholar
* Rannala B, Reeve JP : High-resolution multipoint linkage-disequilibrium mapping in the context of a human genome sequence. _Am J Hum Genet_ 2001; 69: 159–178. Article CAS Google Scholar
* Rossi S, Testa F, Li A _et al_: Clinical and genetic features in Italian Bietti crystalline dystrophy patients. _Br J Ophthalmol_ 2013; 97: 174–179. Article Google Scholar * Halford S,
Liew G, Mackay DS _et al_: Detailed phenotypic and genotypic characterization of bietti crystalline dystrophy. _Ophthalmology_ 2014; 121: 1174–1184. Article Google Scholar * Wang F, Wang
H, Tuan HF _et al_: Next generation sequencing-based molecular diagnosis of retinitis pigmentosa: identification of a novel genotype-phenotype correlation and clinical refinements. _Hum
Genet_ 2014; 133: 331–345. Article CAS Google Scholar * Haddad NM, Waked N, Bejjani R _et al_: Clinical and molecular findings in three Lebanese families with Bietti crystalline
dystrophy: report on a novel mutation. _Mol Vis_ 2012; 18: 1182–1188. CAS PubMed PubMed Central Google Scholar Download references ACKNOWLEDGEMENTS We would like to thank the families
and patients who participated in this work. In addition, we are grateful to NEI project EYE000272 for support. MBG has received support from the Department of Ophthalmology at UCLA, the
Harold and Pauline Price Foundation, and from Research to Prevent Blindness, New York, NY. ZBJ has received support from the National Natural Science Foundation of China (81522014,
81371059). AI is supported by the Research to Prevent Blindness, New York, NY (Physician Scientist Award to AI and unrestricted grant to the UTHSC Department of Ophthalmology). AUTHOR
INFORMATION AUTHORS AND AFFILIATIONS * Ophthalmic Genetics and Visual Function Branch, National Eye Institute, Bethesda, MD, USA Xiaodong Jiao, Anren Li, Xinjing Wang & J Fielding
Hejtmancik * Division of Ophthalmic Genetics, The Eye Hospital of Wenzhou Medical University, China Zi-Bing Jin * Department of Ophthalmology, Center for Retinal Degenerations and Ophthalmic
Genetic Diseases, Duke Eye Center, Duke University Medical Center, Durham, NC, USA Alessandro Iannaccone * Center for Genetic Eye Diseases and Department of Ophthalmic Research, Cole Eye
Institute, Cleveland Clinic Foundation, Cleveland, OH, USA Elias I Traboulsi * Stein Eye Institute and the UCLA Departments of Ophthalmology and Human Genetics, Los Angeles, CA, USA Michael
B Gorin * Eye Clinic, University of Campania L Vanvitelli, Caserta CE, Italy Francesca Simonelli Authors * Xiaodong Jiao View author publications You can also search for this author inPubMed
Google Scholar * Anren Li View author publications You can also search for this author inPubMed Google Scholar * Zi-Bing Jin View author publications You can also search for this author
inPubMed Google Scholar * Xinjing Wang View author publications You can also search for this author inPubMed Google Scholar * Alessandro Iannaccone View author publications You can also
search for this author inPubMed Google Scholar * Elias I Traboulsi View author publications You can also search for this author inPubMed Google Scholar * Michael B Gorin View author
publications You can also search for this author inPubMed Google Scholar * Francesca Simonelli View author publications You can also search for this author inPubMed Google Scholar * J
Fielding Hejtmancik View author publications You can also search for this author inPubMed Google Scholar CORRESPONDING AUTHOR Correspondence to J Fielding Hejtmancik. ETHICS DECLARATIONS
COMPETING INTERESTS The authors declare no conflict of interest. ADDITIONAL INFORMATION Supplementary Information accompanies this paper on European Journal of Human Genetics website
SUPPLEMENTARY INFORMATION SUPPLEMENTARY TABLE S1 (XLSX 10 KB) RIGHTS AND PERMISSIONS Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Jiao, X., Li, A., Jin, ZB. _et al._
Identification and population history of CYP4V2 mutations in patients with Bietti crystalline corneoretinal dystrophy. _Eur J Hum Genet_ 25, 461–471 (2017).
https://doi.org/10.1038/ejhg.2016.184 Download citation * Received: 10 June 2016 * Revised: 15 November 2016 * Accepted: 24 November 2016 * Published: 04 January 2017 * Issue Date: April
2017 * DOI: https://doi.org/10.1038/ejhg.2016.184 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is
not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative