- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
A 6kb rDNA probe comprising the 18S coding plus spacer sequences has been hybridized to the metaphase chromosomes of apple rootstock cultivar MM106 demonstrating the localization of
ribosomal gene arrays in the vicinity of the telomeric regions of the short arms of chromosomes 6 and 14. The in situ results using digoxygenin labelling coupled to an alkaline phosphatase
immunoassay were confirmed by silver staining for NORs and nucleoli. This study demonstrates the feasibility of molecular cytogenetic analysis of very small chromosomes(1.0–2.7μm) of apple.
Our group is currently developing non-radioactive systems for the in situ hybridization of DNA markers to the chromosomes of apple (Malus sp.) in order to assign marker linkage groups to
physical chromosomes and aid mapping of the apple genome1, 2, 3. Determination of the location of genes and repeat sequences by in situ hybridization of biotinylated probes to metaphase
preparations has been reported in several plant species with larger chromosomes than Malus including Allium4, Pisum5, 6, Crdpis7, Picea8 and Triticum9, 10, 11. However, there has been
limited work so far on plant species with very small chromosomes where cytogenetic analysis is more diffcult. There are two recent reports on karyotype analysis for Arabidopsis which has
chromosomes of comparable size but fewer in number (2n=10) than Malus (2n=34)12, 13. This group used digoxygenin labelling coupled with a fluorescent detection system to determine the
chromosomal localization of rDNA (9kb probel and of a smaller tandem repeat sequence(380bp probe). Although there is a recent report8 of successful in situ hybridization to chromosomes of a
woody gymnosperm (Picea), there are no such reports for woody angiosperms.
This paper describes the use of digoxygenin-labelled apple rDNA sequences(18S coding sequence plus spacer)as a probe along with an enzyme—linked colour reaction to hybridize to MM106
chromosomes. The position of the nucleolar organiser regions, the site of ribosomal RNA transcription14, 15 was determined by silver staining to provide independent verification of the rDNA
probe results. The number of active ribosomal gene loci can also be confirmed by counting nucleoli (sites of ribosome biogenesis)in interphase nuclei13, 14, 16, 17, 18.
Actively growing root tips were harvested from glasshouse-grown cuttings of the apple rootstock 'MM106' 'Northern Spy' (Malus × domestica Borkh.) × Malling I-derived from M. paradisia19,
Root tips were pretreated with a saturated paradichlorobenzene solution for 5-6 h at room temperature, rinsed in distilled water f or 5 min and fixed in freshly prepared acidic acid /
ethanol (1:3) at 4°C overnight. Cell walls were hydrolysed by incubation in a solution of 2%cellulase and 2%pectinase(Calbiochem-Behring)at pH 4. 5 for 90-120 min at 37°C. Roots were washed
2-3 min in distilled water and transferred to 45%acetic acid before scraping out, macerating. and squashing the meristematic cells. Preparations were frozen in liquid air, coverslips were
removed and slides were immersed in 95%ethanol for 30-60 min before air drying and then stored at-70°C. Following Giemsa staining20, mitotic spreads with cleanly separated chromosomes were
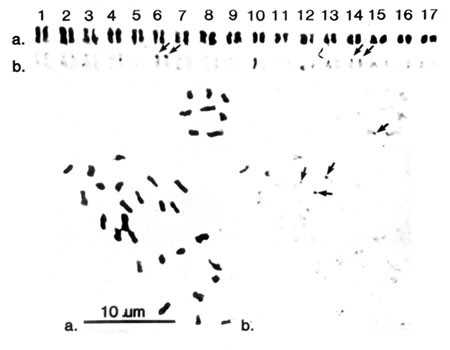
identified, photographed under an on objective and karyotypes prepared (Fig 1A).
Root tip metaphase chromosome spread of apple rootstock 'MM106' with karyotype.
b. In situ hybridization with an 18S plus spacer rDNA probe detected with the alkaline phosphatase-linked immunoassay. Sites are indicated by arrows.
The rDNA used as a probe (a gift from Dr. C J Simon)was a 6kb rDNA fragment from genomic DNA isolated from the crab apple 'Whire Angel' cloned into pUCl3. It included an 18S coding region
and an intergenic spacer21. The fragment was excised with Eco RI and then labelled with digoxygenin-dUTP by random priming according to the manufacturer's instructions (Boehringer Mannheim
GmBh).
Prior to hybridisation, metaphase chromosome preparations were destained in acetic acid ethanol (1:3) for approximately 5-10 mih, transferred to 95% ethanol, and air dried. The slides were
treated with DNase-free RNase f(100μg/ml in 2 × SSC) for 60 min at 37°C, rinsed briefly in 2 × SSC at room temperature, and incubated for 30 min in 2 × SSC at 70°C before dehydration through
70%, 95% and 100% ethano1. Slides were then air dried.
Twenty-five μl of hybridisation mixture (20-40 ng of digoxygenin labelled probe in 6 × SSC, 45% formamide and 5 × Denhardt's solution) was placed on each side, covered with a siliconized
coverslip (22 × 22 mm) and sealed with a fast-drying glue. Denaturation was performed by placing slides on a hot plate at 84.5-85 °C (fmonitored by a thermopile) for 7-10 min and quenching
was carried out on ice for 5 min. Hybridization was performed overnight in a humidity chamber at 40°C. After removing the coverslips, slides were washed thoroughly three times with 2 × SSC
at room temperature for 5 min, with PBS and 0.1% Triton X-100 for 3 rain and then again three times in PBS.
Controls included either a preparation pretreated with 50μg/ml DNase for 30 min at 37°C or a normal preparation carried through the hybridization procedure but without a probe.
Detection of sites of hybridization of the digoxygenin-labelled probe with an antibody-enzyme conjugate (anti-digoxygenin-alkaline phosphatase) and its visualisation by an enzyme-linked
colour reaction (substrates BCIP/NBT) was according to the manufacturer's instructions (Boehringer Mannheim GmbH). The preparations were photographed and the sites of hybridization assigned
to specific chromosomes according to the Giemsa karyotype.
Giemsa-stained metaphase chromosome preparations were first destained, and then re-stained with silver nitrate according to either one of two equally successful methods. Following the
procedure of Lacadena et a1.17, 3-4 drops of freshly prepared 0.7% (w/v) aqueous silver nitrate(adjusted to pH 3.0 with formic acid) on chromosome preparations, spread on slildes covered
with coverslips and incubated overnight in a humidity chamber at 50°C . The other technique involved the one-step colloidal silver method as described by Howell and Black22. Sites of intense
black staining (NORs) were assigned to chromosomes. Sites of brown or diffuse staining were non-specific in nature and were ignored.
Measurements were made from photographs of eighteen metaphase spreads for all three types of staining and relative length, arm ratio, were calculated according to Macgregor and Varley20.
Interphase cells from root tips were stained by the aqueous silver nitrate/colloidal developer method of Howell and Black22.
Analysis of eighteen metaphase spreads stained by Giemsa, silver staining, and in situ hybridization confirmed that 'MM106' is a diploid (2n=34) and that its chromosomes can be arranged in
seventeen pairs (Figs 1 and 2) 23. The measurements of relative chromosome length, and arm ratio indicated that a11 the chromosomes are submetacentric (Tab 1). Their size ranged from
1.0–2.7μm, which is slightly larger than reported earlier (0.5–1.0 μm) by Lespinasse et a1.23. Secondary constrictions which have been associated with plant NORs in some studies 4, 5, 10,
16, 24 were not observed.
Silver staining of root tip metaphase chromosomes of apple rootstock 'MM106'. Arrows indicate NORs on spread and karyotype. Bar: 10μm
Hybridization of the rDNA sequence was detected at a 90% frequency(over ten preparations) as patches of dark purplish-brown precipitate at the end of the short arms of chromosomes 6 and 14
(Fig 1b), while control preparations(either DNase pretreatment of chromosomes or hybridization without a probe) failed to give such a colour reaction.
Nucleolar organiser regions stained with silver were visible as tight black spots whereas non-specific staining was yellowish brown and tended to be more diffuse. The ten preparations
examined indicated the presence of NORs at a frequency of 70% for chromosome 6 and 90% for chromosome 14. NORs were present at the end of the short arms for both chromosome pairs. These
results confirm the localization of ribosomal RNA genes detected with in situ hybridisation by the rDNA probe (Fig 2) since the acidic silver staining of ribonucleic protein accumulating
around active NORs in interphase and remaining at mitosis is an indication of gene activity at rDNA sites during the preceding interphase14.
When silver stained interphase preparations were screened to determine the maximum number of nucleoli and hence number of active rDNA loci, most interphase nuclei were found to contain one
large nucleolus formed by complete fusion of nucleoli during the cell cycle, but few contained two, three, or the primary number of four25, indicating that all four rDNA loci can be active
as reported for AraNdopsis13. The nucleoli were of two size classes (Fig 3), reflecting differences in amount or rate of transcription at the rDNA loci on chromosome pairs 6 and 14, or
difierent rates of assembly or export of ribosomes26.
Silver-stained interphase nucleus of apple rootstock 'MM106' showing 4 nucleoli.
Onr finding that rDNA genes in Malus are localized on two pairs of chromosomes is consistent with earlier in situ hybridization for the diploid angiosperm species including A11ium cepa, A.
fistulogum, AraNdopsis thaliana, Pisum sativum, and Hordeum vulgare. In contrast in Secale cereale hybridization only one pair of chromosomes has been reported 4, 5, 13, 24.
The combination of in situ hybridization of metaphase preparations with a homologous rDNA probe with acidic silver staining of NORs and nucleoli has enabled the assignment of rDNA genes to
the telomeric regions of the short arms of chromosomes 6 and 14 in the apple rootstock 'MM106'. The digoxygenin labelling of the probe coupled with an antibody-alkaline phosphatase colour
detection system proved to be a sensitive, reliable and rapid method for the identification of sites of hybridization to small chromosomes (1.0–2.7 μm) of this woody perennial.
Future studies for in sjtu hybridization to apple chromosomes will ultimately relate information from linkage maps to the physical chromosomes.
This research was funded by the New Zealand Foundation for Research, Science and Technology, Contract Number C06209.
The Horticulture and Food Research Institute of New Zealand Limited,
Mt Albert Research Centre, Private Bag 92 169, Auckland, New Zealand
Batchelar Research Centre, Private Bag 11 030, Palmerston North, New Zealand
Anyone you share the following link with will be able to read this content:









